Fossils have fascinated mankind for millennia. They provide clues about the great intrigue of the past history of life. Unfortunately some fossils are not well preserved, and some things we call fossils are not fossils at all. Our fascination with fossils and what they represent can cause us to sometimes "see" what we want to see instead of what is really there. Some of the most intense scientific battles have been about the proper identification of objects which some consider to be genuine fossils and others consider to be false fossils. Examples of false fossils warn us to be cautious, especially when dealing with ill defined objects which, in spite of varied claims, may or may not be real fossils.
A Case History
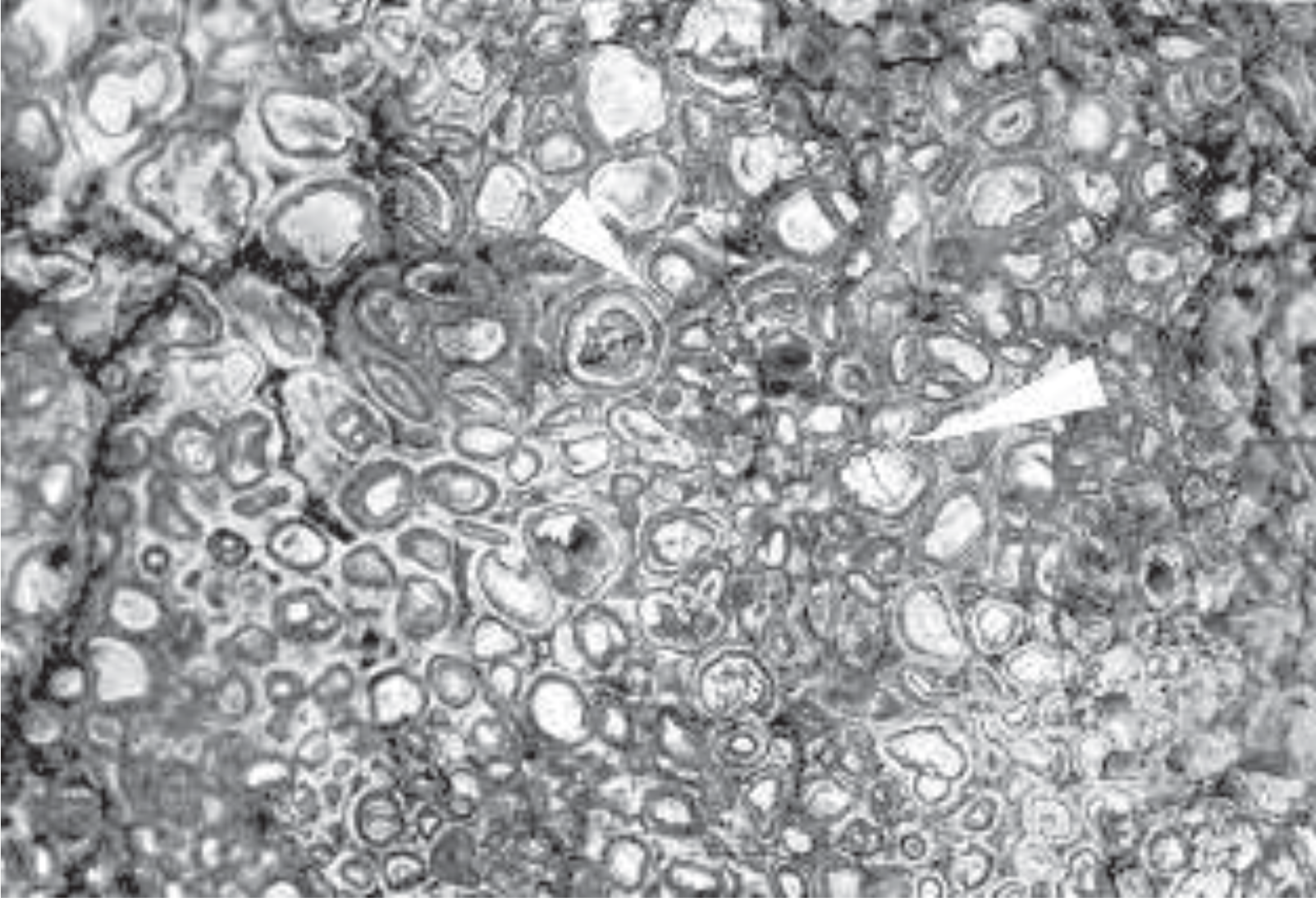
Just east of the gigantic world-famous Carlsbad Caverns in New Mexico, are some intriguing limestone layers consisting of closely packed pea-size spheres (Figure 1). How did these form? There are several ideas, and controversy over their origin has been smoldering for most of this century. The layers of rocks, which at a distance look very ordinary, immediately capture your attention when you get close and notice millions of small marble-like spheres. They are called pisoliths. The term comes from the Greek word "pisos," meaning pea. A rock consisting of pisoliths is called a pisolite.
FIGURE 1. Life-size figure of the weathered surface of a pisolite from the Permian Yates Formation in Walnut Canyon near Carlsbad, New Mexico. The individual spheres are called pisoliths. The short arrow points to an example of polygonal fitting; the long arrow points to concentric layers going around two pisoliths, indicating growth after the appearance of the original pisoliths.
The conventional wisdom during the early part of the century was that these pisoliths were formed by the action of algae growing over the surface of fine grains. The grains became larger as the algae facilitated chemical precipitation of lime (calcium carbonate) and/or the capture of fine sediments. As the grains were rolled around by moving water, growth would take place on all sides producing a somewhat spherical pisolith.
In 1929 the state paleontologist of New York, Rudolf Ruedemann,[1] emphasized the algal origin of these spheres, but a dozen years later two other scientists would disagree. J. V. Pia[2] reported that he could not find any algae, while Walter Lang[3] reported on a few algae, and questioned their significance in producing the pisoliths. At the same time Harlan Johnson[4] reported that he could not find any algal cell structure in the pisoliths, but he believed the majority of them were formed by the action of algae. About the middle of the century, a group of geologists (Newell, Rigby, Fischer, Whiteman, Hickox, and Bradley) published a book[5] on the geology of the region in which they discussed the origin of the pisoliths. While they discussed a number of reasons why algae could not have produced them, they ended up siding with the prevailing view that they had been produced by algae.
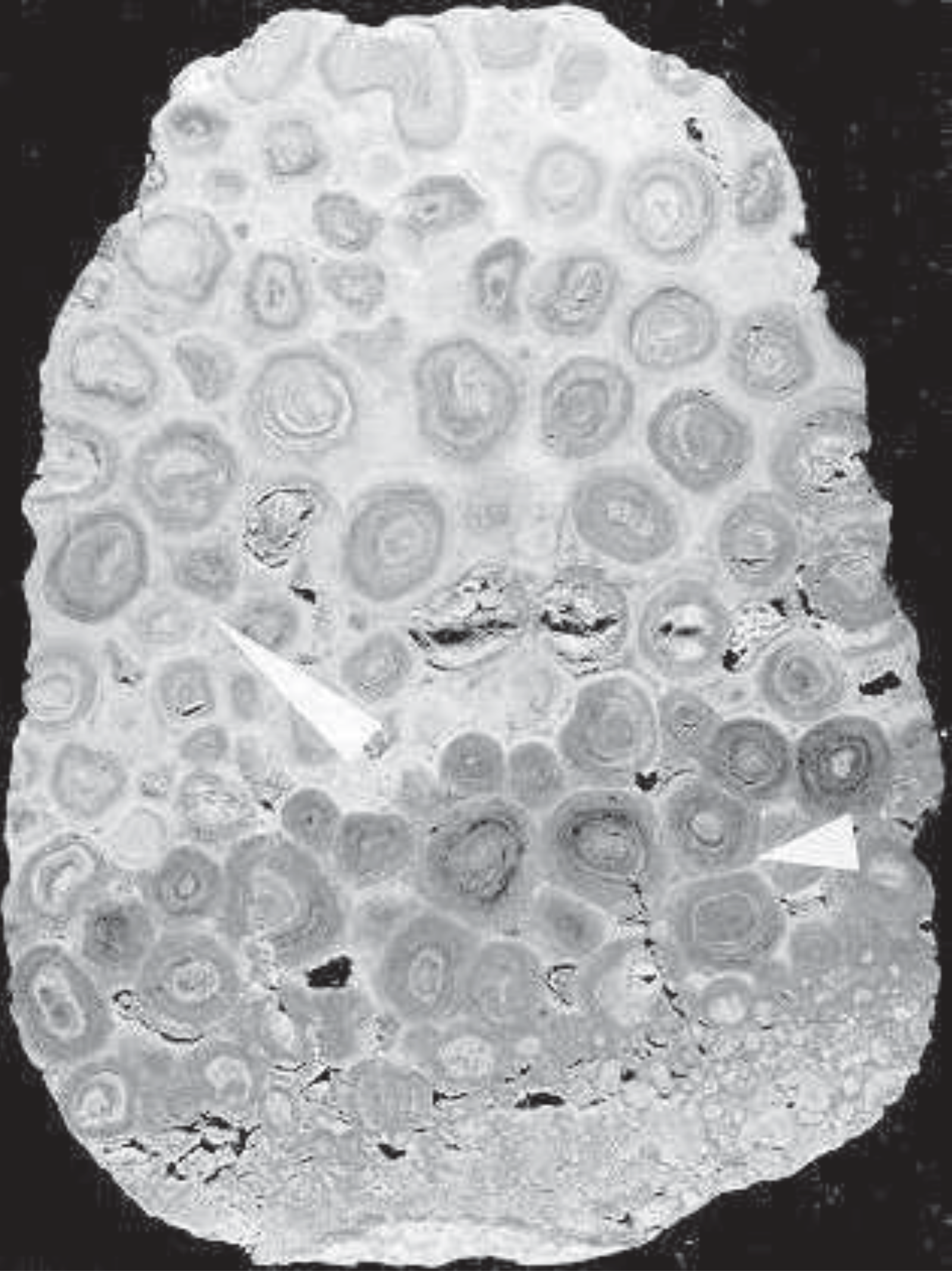
The most dramatic change in thinking about the origin of these spheres took place just a few years later when two investigators, Robert Dunham[6] and Carroll Thomas,[7] working independently, concluded that the pisoliths were not the result of the work of algae, but were formed inorganically, underground, by the gradual accumulation of their many lime layers (Figure 2) around an original nucleus. As water occasionally percolated down through the normally dry soil of the region it facilitated the replacement of the original lime sediments with layers of denser concentration which form the pisoliths. The common spherical concretions we find in many sedimentary rocks are thought to have formed in a similar way. In the region of Carlsbad Caverns where there are many limestone caves lined on the inside with millions of alluring stalactites and stalagmites which are formed from water transported lime, Dunham's and Thomas' novel model is not so hard to imagine. The common formation in soil of a hardpan layer below the surface illustrates how minerals can be easily transported underground by water. Some of the evidence presented by Dunham and Thomas includes: (1) Reverse graded bedding with the larger pisoliths on top of smaller ones (Figure 2). Normally in transported sediments it is the opposite, with the larger particles below. (2) The fitting of the pisoliths against each other (polygonal fitting) as though they grew next to each other (short arrows, Figures 1, 2). (3) Layers enclosing several pisoliths (long arrows, Figures 1, 2). These seem to unequivocally demonstrate that at least some of the lime layers are produced underground. Their formation would have to follow any process of development or emplacement of the spherical bodies. (4) The absence of algae. Algae which require light for growth would be essentially absent below the surface of the ground where Dunham and Thomas proposed the pisolites developed.
FIGURE 2. Polished slab of a pisolite. From the Permian Yates Formation in Walnut Canyon, near Carlsbad, New Mexico. Note the smaller pisoliths at the bottom (reverse graded bedding), the short arrow pointing to polygonal fitting of the pisoliths, and the long arrow pointing to lamina that surround more than one pisolith. The rock is 12 cm in length.
Soon some objected to the model. After giving due consideration to various possibilities, C. G. St. C. Kendall[8] opted for a composite origin involving both algae and inorganic precipitation. The most severe challenge came from two geologists, Mateo Esteban and Lloyd Pray[9] who strongly disagreed with the model and vaguely suggested some kind of formation in water and also just below the water-sediment line in a hypersaline environment. They did not suggest that algae were involved.
Despite the suggestions by Esteban and Pray, the model of underground formation, in the vadose region above the water table, has gained widespread approval.[10] It was soon applied to many similar deposits in neighboring Texas,[11] as well as other parts of the world including Italy, Canada and Morocco.[12] Recently the underground precipitation model has received further support by experiments that produce similar structures in unconsolidated mud.[13] While not all are agreed, the strong prevailing opinion is that the pisoliths are not of algal origin.
What is a Fossil?
The question of whether the pisoliths mentioned above are produced by algae or by inorganic precipitation determines whether they are fossils. A fossil is any evidence of ancient life. If the pisoliths were not produced by algae, or some other living organisms, they are not fossils. While there is an abundance of thoroughly unquestionable fossils to be found, there are also many problematic forms in the rocks of the crust of the Earth that challenge our innate desire for definitive answers. Sometimes the term pseudofossil is used to designate a form that was thought to be a fossil but that turns out to be of non-living origin.[14] One dictionary describes a pseudofossil as an object mistaken for a fossil by an inexperienced person; however the case of the pisoliths mentioned above does not exonerate experts from the challenge of determining if something is really a fossil. The term dubiofossil is sometimes used when we are more sure that we don't know.
Determination of whether a peculiar form in a rock is a bona fide fossil can, in some cases, be extremely difficult. Examples abound. Preserved mud curls caused by drying have sometimes been interpreted as crab parts; drag marks caused by movement of objects during storms can resemble worm tracks; chemical precipitation, in rose-like shapes, of the mineral pyrite have been interpreted as medusae (jellyfish), as have gas-bubble markings;[15] and some supposed sponge-like fossil organisms (archeocyathids) have turned out to be forms produced by inorganic crystallization.[16]
The venerable Treatise on Invertebrate Paleontology[17] lists 69 published descriptions of "fossil organisms" originally identified as coral, algae, fungi, sponges, snails, etc., that are most likely of non-biological origin. These misidentified objects appear to have been produced by unusual depositional conditions. Brooksella canyonensis is a "fossil" which resembles a star-shaped crack. It has an impressive pedigree of interpretations, including: (1) the body fossil of a jellyfish, (2) the reverse imprint of an inorganic fracture system produced by gas evasion, (3) the result of compaction, (4) the imprint of a starlike feeding burrow, or (5) possibly the work of a worm.[18]
Numerous tiny "shells" in the Precambrian of Mongolia caused considerable concern, because they were found in an unexpected location. The discoverers published additional papers supporting their authenticity. But the "shells" turned out to be produced by precipitation of mineral formations around gas bubbles during the preparation of the rock specimens.[19]
In the fossil record we occasionally find what are commonly called "worm tubes." These are elongated tubes of various shapes and orientation. Some are unquestionably genuine fossils, identifiable by the structural patterns left by the organisms that produced them. Others are subject to other interpretations. It is well known that gases and fluids escaping from sediments can form vertical and sinuous tubes. Some horizontal tubes that have been interpreted as being formed by organisms have turned out to be desiccation cracks which later became filled by other sediments.[20]
The problem of pseudofossils is particularly acute in the lowest parts of the geologic record, where evolutionists expect the earliest, simple life forms to have originated. Finding these earliest forms of life has almost become an obsession with some paleontologists. Creationists can interpret lowest fossils as representatives of created forms of microscopic life. Many indications of microscopic life at unexpected depth in rocks have been reported in the professional literature. On the other hand, several investigators have been able to simulate the shape of these presumed simple life forms by inorganic precipitation or by special depositional conditions. Spherical, tube-like, or coiled shapes, characteristic of fossil forms, are easily reproduced from simple inorganic chemicals in the laboratory.[21]
It is to the credit of paleontologists that considerable caution is now being expressed regarding the authenticity of most claims concerning fossils in what is considered to be the oldest sediments, the Archean. In referring to microfossils reported from at least 28 Archean localities, two specialists in this field, William Schopf and Bonnie Packer, state: "However, virtually all have recently been reinterpreted ... as dubiofossils or as nonfossils: pseudofossils, artifacts, or contaminants."[22] Paleontologist Richard Cowen states: "Only a few reports of fossil Archean cells seem to be genuine, out of fifty or more claims."[23] Roger Buick at Harvard refers to a host of problems with the identification of most of these primitive fossils found at North Pole, Australia.[24] (It is called North Pole because, like the real North Pole, it is a notably desolate area.) An old geological dictum stating that "I never would have seen it if I hadn't believed it" seems to apply to many of these cases.
The pseudofossil problem also comes into focus with respect to stromatolites (Figures 3, 4), which are finely laminated sedimentary structures, usually in the centimeter to meter range and often having a mounded or wavy form. Stromatolites are formed underwater, as thin mats of microscopic organisms living on their surface trap or precipitate minerals, which are then incorporated into a layered structure. There is a question as to whether what appears to be a fossil stromatolite may have formed biologically, or whether it is just the passive accumulation of fine layers of sediment which has been subjected to deformation. In the latter case they would not be fossils. The sedimentologist Robert Ginsburg points out that "Almost everything about stromatolites has been, and remains to varying degrees, controversial."[25] Stromatolite specialist Paul Hoffman notes: "Something that haunts geologists working on ancient stromatolites is the thought that they might not be biogenic at all."[26] The well-known paleontologist Charles Walcott, who for twenty years was Director of the US Smithsonian Institution, described 5 new genera and 8 new species of stromatolites believed to be of biological origin. Each of these has since been reinterpreted as inorganic by at least one investigator.[27] Even presently forming "stromatolites" can be enigmatic. A number of "stromatolites" described in various parts of Scandinavia have been reinterpreted more recently as being of non-biological origin;[28] however, there are many unquestionable living stromatolites over Earth's surface.
FIGURE 3. View of Precambrian rocks of the Chuar Group near Kwagut Butte in the Grand Canyon of the Colorado River, Arizona. The mounded rocks scattered in the vegetation are interpreted as stromatolites. The arrow points to one of them.FIGURE 4. Cross-section of a stromatolite from the region of Figure 3. Note the wavy layers in the stromatolite. The specimen is about 40 cm across.
Stromatolites are also found in the deep rocks, and their interpretation is more equivocal from both the creationist and evolutionist perspectives. Stromatolites are an important part of the evolutionary scenario of early life; but like many of the fossils in the deeper rocks, their identification is problematic. Some widely accepted examples of ancient stromatolites have been reinterpreted as precipitation and soft sediment deformation.[29] Paleobotanist A. H. Knoll of Harvard points out: "no Early Archaean stromatolites are known to contain micro-fossils. Thus, abiological alternatives must be considered."[30]
The correct identification of fossil stromatolites in deep rocks is important to the question of the origin of life. Estimates of the age of these fossils is complicated by the recent discovery of living stromatolites actively forming in rock cavities such as are sometimes found in coral reefs. These deposits are called endostromatolites. Sediment accumulation on an endostromatolite is facilitated by bacteria that do not require light as an energy source. Claude Monty, a biosedimentologist from the University of Liège in Belgium, suggests that endostromatolites can form in rock cavities at depths of at least 3 km.[31] This raises the question as to whether some stromatolites in the deeper rocks, possibly growing in caves, might be endostromatolites of recent origin. The status of our knowledge regarding these ancient, or assumed-ancient, stromatolites is inadequate, and firm conclusions cannot be drawn.
Life on Mars
The planet Mars has several similarities with Earth. It has been considered as the most likely place beyond Earth where life could exist in our solar system. Occasionally we even fantasize about little green men from Mars. Evolutionists sometimes wonder if life could have evolved independently on our close planetary neighbor, and some creationists wonder if life could have been created there. The extent of life throughout the Universe is one of our most profound questions.
In 1884 the French astronomer E. L. Trouvelot thought that he could see slow color changes in some of the grayish areas of Mars, thus implying seasonal changes in the Martian vegetation. Three years later the Italian astronomer Giovanni Schiaparelli noted long lines on the surface of Mars which he called "canali." Early this century the American astronomer Percival Lowell pursued the same theme and thought he observed a network of canals on the planet. He reasoned that they were too straight to be of natural causes and suggested that they were built by the inhabitants of Mars to bring water from the poles to their crops near the arid equator. Such speculations were laid to rest a few decades ago when the United States and the Soviet Union sent space probes to Mars. They did not find any intelligent life. Highly sensitive detectors found no evidence of life in the Martian soil in the region where the space probes had landed. They did not find any canals either, but instead discovered huge canyons and abundant evidence that huge volumes of water were involved in the sculpturing of parts of the planet. Any suggestion of life on Mars has now been essentially reduced to the possibility of present or past microbial life.
Accounts of fossils purported to represent simple life, that are then disproved by subsequent research, are becoming an old and too often repeated story. The Orgueil Meteorite has been studied for well over a century. The meteorite fell as many fragments in 1864 at Orgueil, France. It contained many carbon compounds which suggested a possible association with living things. This engendered a search for fossil evidence of past life in the meteorite. Several unusual microscopic forms considered to be fossils were found. Their origin became the topic of an unusually lively debate that lasted for years.[32] Some of the fossil evidence turned out to be pollen interpreted by some as recent contamination, and some resembled inorganic structures found in furnace ash. The general conclusion has been to doubt the authenticity of any fossil forms. The Orgueil Meteorite provides little, if any, convincing evidence for the existence of extraterrestrial life.
The search continued. During the late 1960s, when space probes and astronauts were first sent to the Moon, there was initial considerable excitement, followed by disappointment, when it became apparent that there was no life on the Moon. In recent years vast expenditures have been made in listening for radio messages from outer space. Thus far the search for messages from extraterrestrial intelligence has been futile. Much effort has been invested in looking for life beyond Earth. Fossils, dubiofossils, and pseudofossils have played a major role in this search. In several aspects the debate concerning microbial life on Mars has been remarkably similar to the one about the Orgueil Meteorite.
In August 1996 David McKay and his colleagues at the National Aeronautics and Space Administration Johnson Space Center and Stanford University announced that they had found evidence of past life on a meteorite presumed to have come from Mars.[33] The potato-size meteorite is assumed to be around 4.5 billion years old. It supposedly escaped from Mars, and, after an extended sojourn in space, landed on Earth's Antarctic ice an estimated 13,000 years ago. This meteorite, which is designated as ALH84001, is thought to have come from Mars because it is similar to other meteorites that have been found in the same Antarctic region and have gas bubbles whose composition is similar to the atmosphere of Mars, and not of Earth. There is little disagreement, at least within the planetary scientific community, about the origin of this meteorite. But there is considerable debate about the evidence for life in ALH84001. The opposition began immediately after the announcement of its discovery.[34] Arguments in favor of living forms include the presence of organic compounds, magnetite crystals thought to have been produced by microorganisms, and carbonate minerals which are often associated with life. All of these arguments and others have been challenged, either as not representing life, or as indicating contamination from Earth's environment.
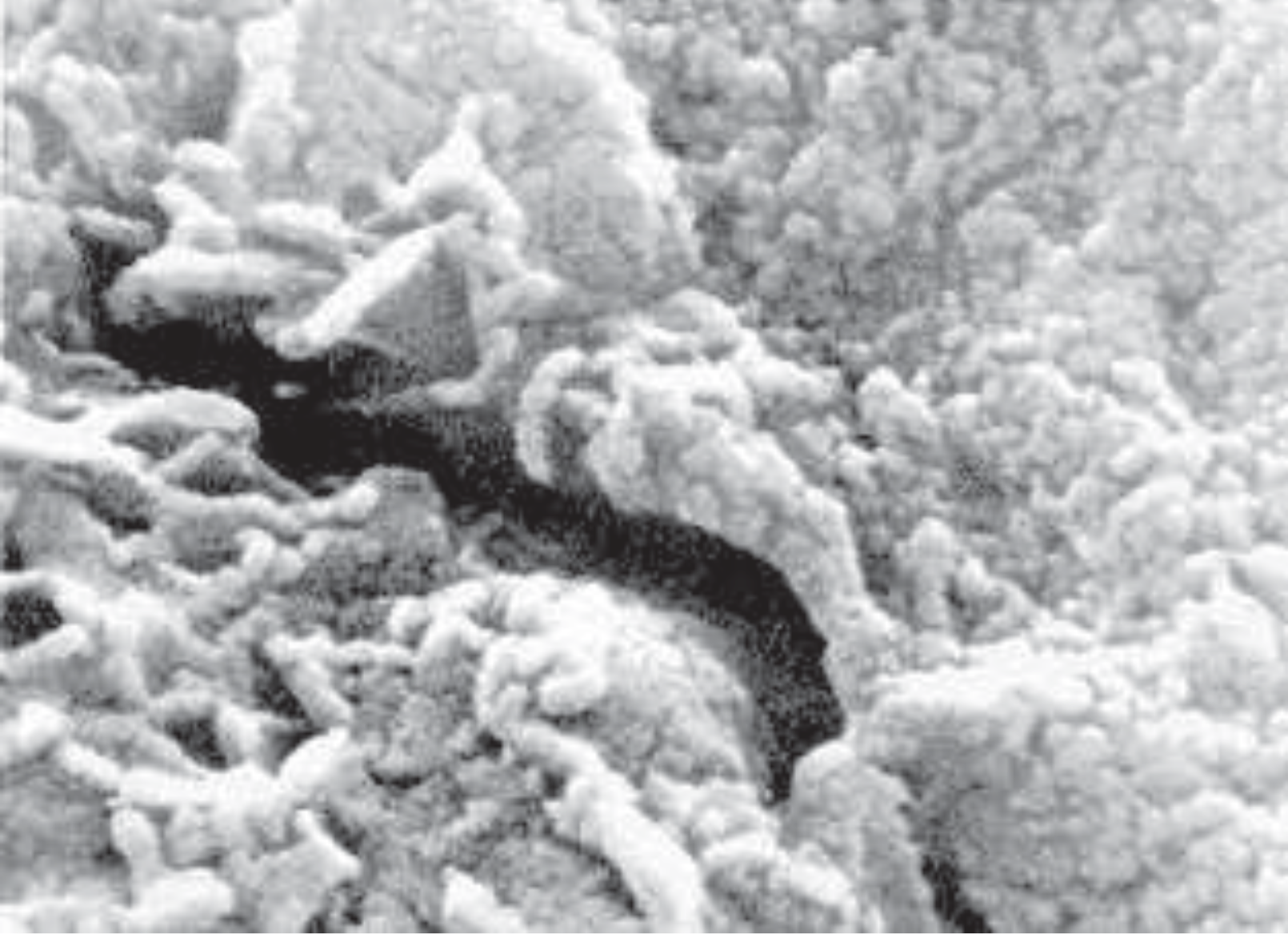
Perhaps the most interesting suggestion by McKay and his group was that tiny filaments and ovoid microscopic structures found in the carbonate minerals of ALH48001 represent fossils (Figures 5 and 6). If it could be demonstrated that these are genuine fossils, much of the rest of the debate would be irrelevant. Are these elongated forms evidence of past life, or do they represent pseudofossils. The so-called fossils found have a slight resemblance to bacteria, but it has been pointed out that the fossils are too small to represent normal bacteria. On the other hand, it is argued that there are organisms on Earth much smaller than ordinary bacteria. Others point out that the so-called worm-like fossils (Figures 5, 6) could be produced by crystalline growth on the edge of layers, or they may be just the protruding edges of mineral layers. They also could be only artifacts resulting from the complex specimen preparation process. In referring to the evidence for life in AHL84001, planetary scientist John Kerridge of the University of California at San Diego states "Now I think they don't have a shred of evidence to back it up."[35] The last chapter about life on Mars has not yet been written. An ambitious program giving further study to the planet is under way. Final conclusions should not be drawn until we have more data, but the present fossil evidence is highly dubious.
FIGURE 5. High-resolution-scanning electron microscope image from the meteorite ALH48001. The elongated tube-like form in the middle is less than 1/100th the width of a human hair. It is considered to be a prime example of a possible fossil from Mars. Photo courtesy NASA.FIGURE 6. Electron microscope image of possible bacterial-like organisms thought to have come from Mars. The fine filamentous structures, which are less than 1/100th the diameter of a human hair, were found in carbonate minerals from the meteorite ALH48001. Photo courtesy NASA.
Conclusions
There are many good fossils to be found around the world. There are also many questionable ones. Unfortunately there are too many genuine pseudofossils. Determining whether a particular form is a genuine fossil is not always easy; it can also be a problem to identify a genuine pseudofossil. Our desires to discover and to proclaim our discoveries can lead us into strange pathways where the questionable can become very real to us. The long list of pseudofossils now bedecking the scientific literature should warn us to be cautious about any unwarranted claims about fossils. In the realm of poorly defined structures that look like fossils, reserving judgment is a virtue.
ENDNOTES
[1]Ruedemann R. 1929. Coralline algae, Guadalupe Mountains. American Association of Petroleum Geologists Bulletin 13:1079-1080.
[2]Pia JV. 1940. As quoted in: King PB. 1948. Geology of the Southern Guadalupe Mountains Texas. U.S. Geological Survey Professional Paper 215:79-80.
[3]Lang WB. 1937. The Permian Formations of the Pecos Valley of New Mexico and Texas. American Association of Petroleum Geologists 21:833-898.
[4]Johnson JH. 1942. Permian lime-secreting algae from the Guadalupe Mountains, New Mexico. Geological Society of America Bulletin 53:195-226.
[5]Newell ND, Rigby JK, Fischer AG, Whiteman AJ, Hickox JE, Bradley JS. 1953. The Permian Reef complex of the Guadalupe Mountains region, Texas and New Mexico: a study in paleoecology. NY. Hafner Publishing Co.
[6]Dunham RJ. 1972. Capitan Reef, New Mexico and Texas: facts and questions to aid interpretation and group discussion. Society of Economic Paleontologists and Mineralogists, Permian Basin Section, Publication 72-14.
[7]Thomas CM. 1968. Vadose pisolites in the Guadalupe and Apache Mountains, West Texas. in: Silver BA, editor. Guadalupian Facies, Apache Mountains area, West Texas. Symposium and Guidebook, 1968 Field Trip. Society of Economic Paleontologists and Mineralogists, Permian Basin Section, Publication 68-11:32-48.
[8]Kendall CGSC. 1969. An environmental re-interpretation of the Permian evaporite/carbonate shelf sediments of the Guadalupe Mountains. Geological Society of America Bulletin 80:2503-2526.
[9](a) Esteban M. 1982. Comments on 'Petrology of recent caliche, pisolites, spherulites, and speleothern deposits from central Texas' by H.S. Chafetz and J.C. Butler. Sedimentology 29:441-445; (b) Esteban M. 1976. Vadose pisolite and caliche. American Association of Petroleum Geologists 60:2048-2057; (c) Esteban M, Pray LC. 1975. Subaqueous, syn-depositional growth of in-place pisolite, Capitan Reef Complex (Permian), Guadalupe Mountains, New Mexico and West Texas. Geological Society of America Abstracts with Programs 7(7):1068-1069; (d) Esteban M, Pray LC. 1977. Origin of the pisolite facies of the shelf crest. In: Upper Guadalupian facies Permian Reef complex, Guadalupe Mountains, New Mexico and West Texas. 1977 Field Conference Guidebook. Society of Economic Paleontologists and Mineralogists, Permian Basin Section, Publication 77-16: 479-483.
[10](a) Jacka AD, Thomas CM, Beck RH, Williams KW, Harrison SC. 1972. Guadalupian depositional cycles of the Delaware Basin and Northwest Shelf. In: Elam JG, Chuber S, editors. Cyclic sedimentation in the Permian Basin. 2nd ed. Midland, TX West Texas Geological Society, p 151-195; (b) Smith DB. 1974. Sedimentation of upper Artesia (Guadalupian) cyclic shelf deposits of northern Guadalupe Mountains, New Mexico. American Association of Petroleum Geologists Bulletin 58:1699-1730.
[11](a) Chafetz HS, ButlerJC. 1980. Petrology of recent caliche pisolites, spherulites, and speleothem deposits from central Texas. Sedimentology 27:497-518; (b) Chafetz HS, Butler JC. 1977. Petrology of recent vadose pisolites, Edwards Formation, Cretaceous of central Texas. Geological Society of America Abstracts with Programs 9(7):924-925; (c) Gibson GG. 1983. Vadose pisolite: additional evidence for a shallow marine to supratidal carbonate origin for the Caballos Novaculite, Trans-Pecos Texas. Geological Society of America Abstracts with Programs 15(2):54.
[13](a) Mazzullo SJ, Birdwell BA. 1989. Syngenetic formation of grainstones and pisolites from fenestral carbonates in peritidal settings. Journal of Sedimentary Petrology 59:605-611; (b) Wright VP. 1990. Discussion: Syngenetic formation of grainstones and pisolites from fenestral carbonates in peritidal settings. Journal of Sedimentary Petrology 60:309-310.
[14]Monroe JS, Dietrich RV. 1990. Pseudofossils. Rocks & Minerals 65(2):150-158.
[15]Cloud P. 1973. Pseudofossils: a plea for caution. Geology 1(3):123-127.
[16]Glaessner MF. 1980. Pseudofossils from the Precambrian, including 'Buschmannia' and 'Praesolenopora.' Geological Magazine 117(2):199-200.
[17]Häntzschel W. 1975. Treatise on invertebrate paleontology, Part W: Miscellanea, Supplement 1. 2nd ed. Boulder, CO: The Geological Society of America; and Lawrence, KS: The University of Kansas, p W169-179.
[19]Li YQ. 1993. China learns cautionary tale from Mongolian 'fossils'. Nature 366:499.
[20]For a discussion and references about "worm tubes," see p 940-942 in: Kauffman EG, Steidtmann JR. 1981. Are these the oldest metazoan trace fossils? Journal of Paleontology 55:923-947.
[21](a) Glaessner ME 1988. Pseudofossils explained as vortex structures in sediments. Senckenbergiana lethaea 69(3/4):275-287; (b) Gutstadt AM. 1975. Pseudo- and dubiofossils from the Newland Limestone (Belt Supergroup, late Precambrian), Montana. Journal of Sedimentary Petrology 45(2):405-414; (c) Jenkins RIF, Plummer PS, Moriarty KC. 1981. Late Precambrian pseudofossils from the Flinders Ranges, South Australia. Transactions of the Royal Society of South Australia 105(2):67-83; (d) Merek EL. 1973. Imaging and life detection. BioScience 23(3):153-159; (e) Pickett J, Scheibnerová V. 1974. The inorganic origin of "anellotubutates." Micropaleontology 20(l):97-102; (f) Service RF 1995. Prompting complex patterns to form themselves. Science 270:1299-1300.
[22]Schopf JW, Packer BM. 1987. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 237:70-73.
[23]Cowen R. 1995. History of life. 2nd ed. Boston, Oxford and London: Blackwell Scientific Publications, p 39.
[24]Buick R. 1990. Microfossil recognition in Archean rocks: an appraisal of spheroids and filaments from a 3500 m.y. old chert-barite unit at North Pole, Western Australia. Palaios 5:441-459.
[25]Ginsburg RN. 1991. Controversies about stromatolites: vices and virtues. In: Müller DW, McKenzie JA, Weissert H, editors. Controversies in modern geology. London, San Diego, and NY. Academic Press, p 25-36.
[26](a) Hoffman P. 1973. Recent and ancient algal stromatolites: seventy years of pedagogic cross-pollination. In: Ginsburg RN, editor. Evolving concepts in sedimentology. The Johns Hopkins University Studies in Geology, No. 21. Baltimore and London: The Johns Hopkins University Press, p 178-191. See also: (b) Grotzinger JP, Rothman DH. 1996. An abiotic model for stromatolite morphogenesis. Nature 383:423-425; (c) Lowe DR. 1994. Abiological origin of described stromatolites older than 3.2 Ga. Geology 22:387-390.
[28]Bjærke T, Dypvik H. 1977. Quaternary 'stromatolitic' limestone of subglacial origin from Scandinavia. Journal of Sedimentary Petrology 47:1321-1327.
[29]Lowe DR. 1994. Abiological origin of described stromatolites older than 3.2 Ga. Geology 22:387-390.
[30]Knoll AH. 1990. Precambrian evolution of prokaryotes and protists. In: Briggs DEG, Crowther PR, editors. Palaeobiology: a synthesis. Oxford and London: Blackwell Scientific Publications, p 9-16.
[31](a) Monty CLV. 1986. Range and significance of cavity-dwelling or endostromatolites. Sediments down-under. Abstracts of the 12th International Sedimentological Congress, Canberra, Australia, p 216; (b) Vachard D, Razgallah S. 1988. Survie des genres Tharama et Renalcis (Epiphytales, a1gues problématiques) dans le Permien supérieur du Djebel Tebaga (Tunisie). Comptes Rendus de L'Académie des Sciences Paris 306(Ser 2):1137-1140.
[32](a) Bernal JD. 1962. Comments. Nature 193:1127-1129; (b) Bernal JD. 1961. Significance of carbonaceous meteorites in theories on the origin of life. Nature 190:129-131; (c) Briggs MH. 1962. Properties of the organic microstructures of some carbonaceous chondrites. Nature 195:1076-1077; (d) Briggs MH, Kitto GB. 1962. Complex organic micro-structures in the Mokoia Meteorite. Nature 193:1126-1127; (e) Claus G, Nagy B. 1961. A microbiological examination of some carbonaceous chondrites. Nature 192:594-596; (f) Fitch F, Schwarcz HP, Anders E. 1962. 'Organized elements' in carbonaceous chondrites; (g) Marnikunian G, Briggs MH. 1963. Some microstructures of complex morphology observed in preparations of carbonaceous chondrites made under sterile conditions. Nature 197:1245-1248; (h) McCall GJH. 1973. Meteorites and their origins. Newton Abbot, Devon: David & Charles; (i) Mueller G. 1962. Interpretation of micro-structures in carbonaceous meteorites. Nature 196:929-932;(j) Mueller G. 1964. 'Impact contamination' of the Mokoia carbonaceous chondrite. Nature 204:567; (k) Nagy B, Claus G, Hennessy DJ. 1962. Organic particles embedded in minerals in the Orgueil and Ivuna carbonaceous chondrites. Nature 193:1129-1133; (l) Pearson R. 1962. Life-forms in carbonaceous chondrites. Nature 194:1064-1065; (m)Urey HC. 1962. Life-forms in meteorites. Nature 193:1119-1123.
[33]McKay DS, Gibson EK, Thomas-Keprta KL, Vali H, Romanek CS, Clemett SJ, Chillier XDF, Maechling CR, Zare RN. 1996. Search for past life on Mars: possible relic biogenic activity in martian meteorite ALH84001. Science 273:924-930.
[34]Anders E; Shearer CK, Papike JJ; Bell JF; Clemett SJ, Zare RN; McKay DS, Thomas-Keprta KL, Romanek CS, Gibson EK, Vali H; Gibson EK, McKay DS, Thomas-Keprta K, Romanek CS. 1996. Evaluating the evidence for past life on Mars. Science 274:2119-2125.
[35]As quoted in: Cowen R. 1996. Searching for life in a Martian meteorite. Science News 150:380, 383.
COVER. Weathered surface of a rock called a pisolite. The photo is from the Permian Yates Formation exposed in Walnut Canyon, near Carlsbad Caverns, New Mexico. The individual spheres forming this rock are called pisoliths. Their origin has been debated for most of this century. Were they formed by algae, or by inorganic precipitation? The photo, taken by the author, is reproduced at about life size.