©Copyright 2018 GEOSCIENCE RESEARCH INSTITUTE
11060 Campus Street • Loma Linda, California 92350 • 909-558-4548
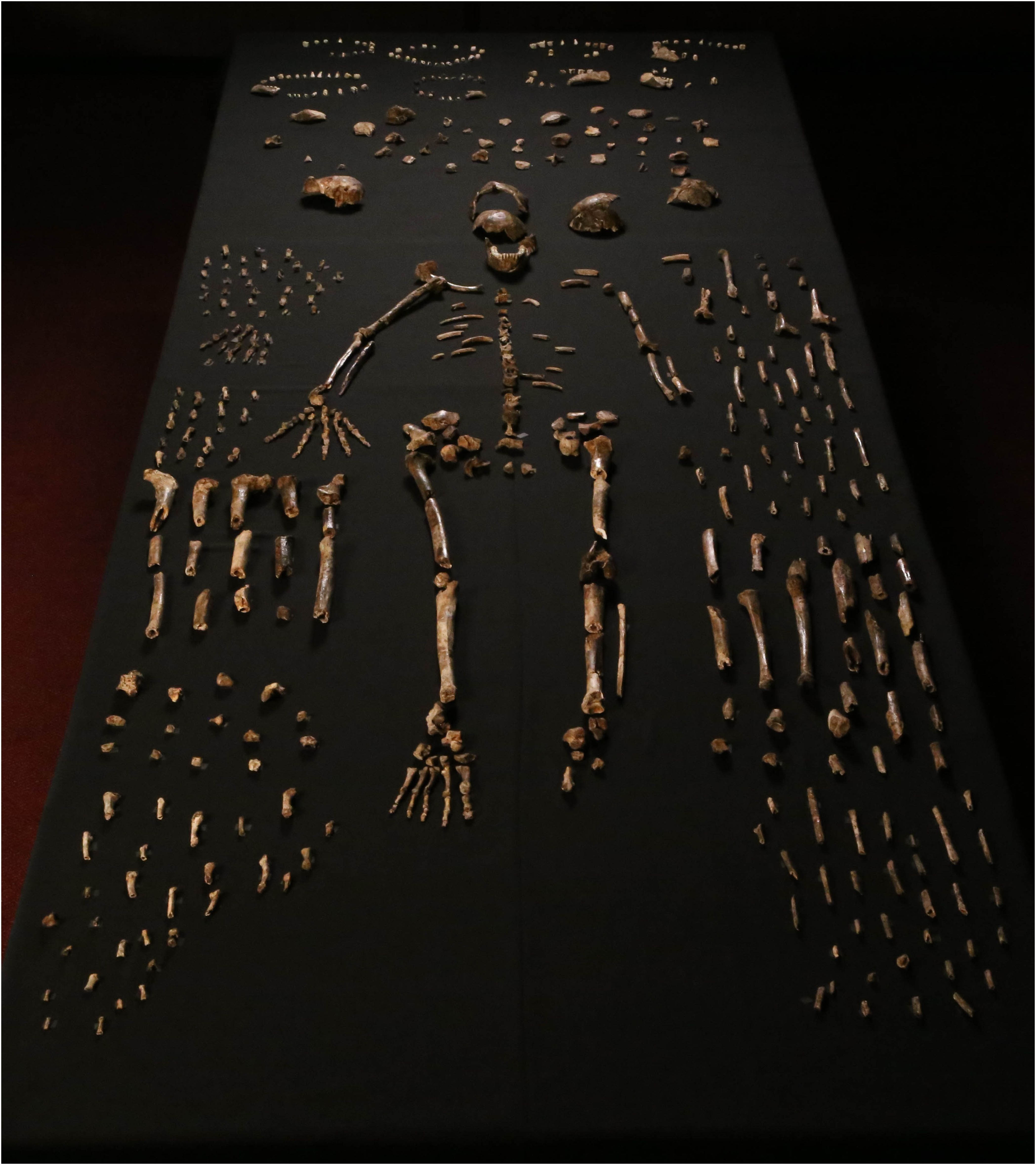
On September 10 2015, an open-access article on the journal eLife (downloadable at http://elifesciences.org/content/4/e09560) presented to the scientific community the newest addition of a species to the hominid fossil record: Homo naledi [1] (Fig. 1). A well-orchestrated unveiling of the discovery included release of an almost 2-hours long documentary produced by PBS and National Geographic entitled “Dawn of Humanity” and the classic amplification by major media outlets of the sensational finding.
See more at: http://elifesciences.org/content/4/e09560#sthash.i...
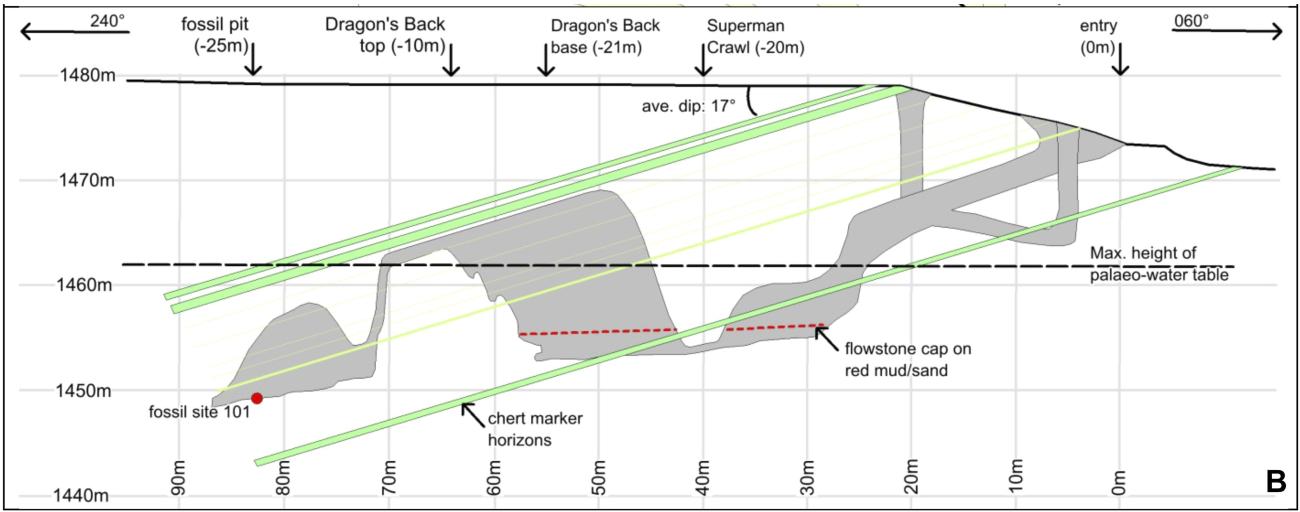
The fossil material described in the paper is indeed remarkable for several reasons. First and foremost, the sheer number of skeletal and dental remains recovered (over 1500, with scores more waiting to be excavated from the same site) makes this “the largest collection of a single species of hominin that has been discovered in Africa so far” [2]. A minimum of 15 individuals is represented in the assemblage, spanning different age groups, from infant to old adult. The remains are fairly well preserved, with some breakage and superficial abrasion, and are generally disassociated except in a few notable cases (e.g., the bones of a hand and those of a lower limb still in articulation) (Fig. 1). The second aspect triggering attention in the news is the mixture of human-like and australopithecine-like anatomical traits exhibited by different recovered parts (e.g., “human-like hands and feet” and “australopith-like pelvis”[3]). Finally, the context in which the fossils are preserved is also extremely intriguing. The remains were found in the top 20 cm of fine sediment covering the floor of a deep chamber in a cave system (called Rising Star) near Johannesburg (South Africa), presently accessible only through narrow passages (down to 20-cm in diameter) with vertical drops of more than 10 m (Fig. 2). It does not appear that the cave chamber had different accesses in the past nor that water transported the remains in the chamber. In addition, the assemblage consists exclusively of hominid remains, with no remains of other animals mixed in (with the exception of the bones of an owl and a few rodent teeth). This setting has led to speculation that the carcasses might have been intentionally disposed in this deep, completely dark recess, as discussed in an accompanying paper on eLife (downloadable at http://elifesciences.org/content/4/e09561) [4].
indicating the site where fossils were found.
http://elifesciences.org/content/4/e09561#F2
For those who treasure knowledge, science, and the Word of God, discoveries like this always elicit an ambivalent reaction. We are excited for a new piece of evidence brought back from the past but saddened in seeing it invariably presented to the masses with an unfiltered endorsement of Darwinian evolution. For example, in the space of a few paragraphs a CNN piece on Homo naledi [5] uses expressions like “new species of human ancestor,” “a new addition to our family tree,” and “new species of human relative.” The implicit message in this use of language is that humans are not the result of an intentional act of divine creation but are just one of the many branches of the tree of universal common descent from ancestral forms. This view contrasts with the alternative model of God creating distinct types (including humans) that might have modified with time. Language is a powerful tool that can be used to advocate specific interpretations of data. Generally, however, data themselves can be compatible with multiple hypotheses. The aim of this article is to use the example of Homo naledi to illustrate the distinction between data and interpretations, and to discuss some of the questions a biblical creationist might have in relation to this new discovery.
Understanding the limitations of data
The Homo naledi fossil assemblage is an excellent example of the limitations intrinsic to the practice of historical sciences. We can only work with what is preserved and available, and build plausible scenarios based on the most logical inference, excluding unsupported hypotheses. Here are a few examples to illustrate the point:
1) Completeness. In terms of quantity of fossil remains and representation of different body parts from one single site, the discovery at Rising Star cave is very impressive. However, even amidst this abundance, the record is still fragmentary. For instance, remains of at least 15 individuals have been recovered so far, but only five partial crania and a few other cranial fragments are represented. Some parts of the cranium are not preserved or are documented only by one specimen. Given these limitations, traits that are presented as diagnostic for the Homo naledi skull are still based on a relatively limited database. The paper specifies that when different specimens preserve the same part of the cranium, they “agree closely in all morphological details”. However, it also mentions exceptions, where differences between crania are interpreted “as related to sex” [6], that is due to variability between male and female individuals. Unfortunately, the paper does not provide a detailed discussion of the hinted differences.
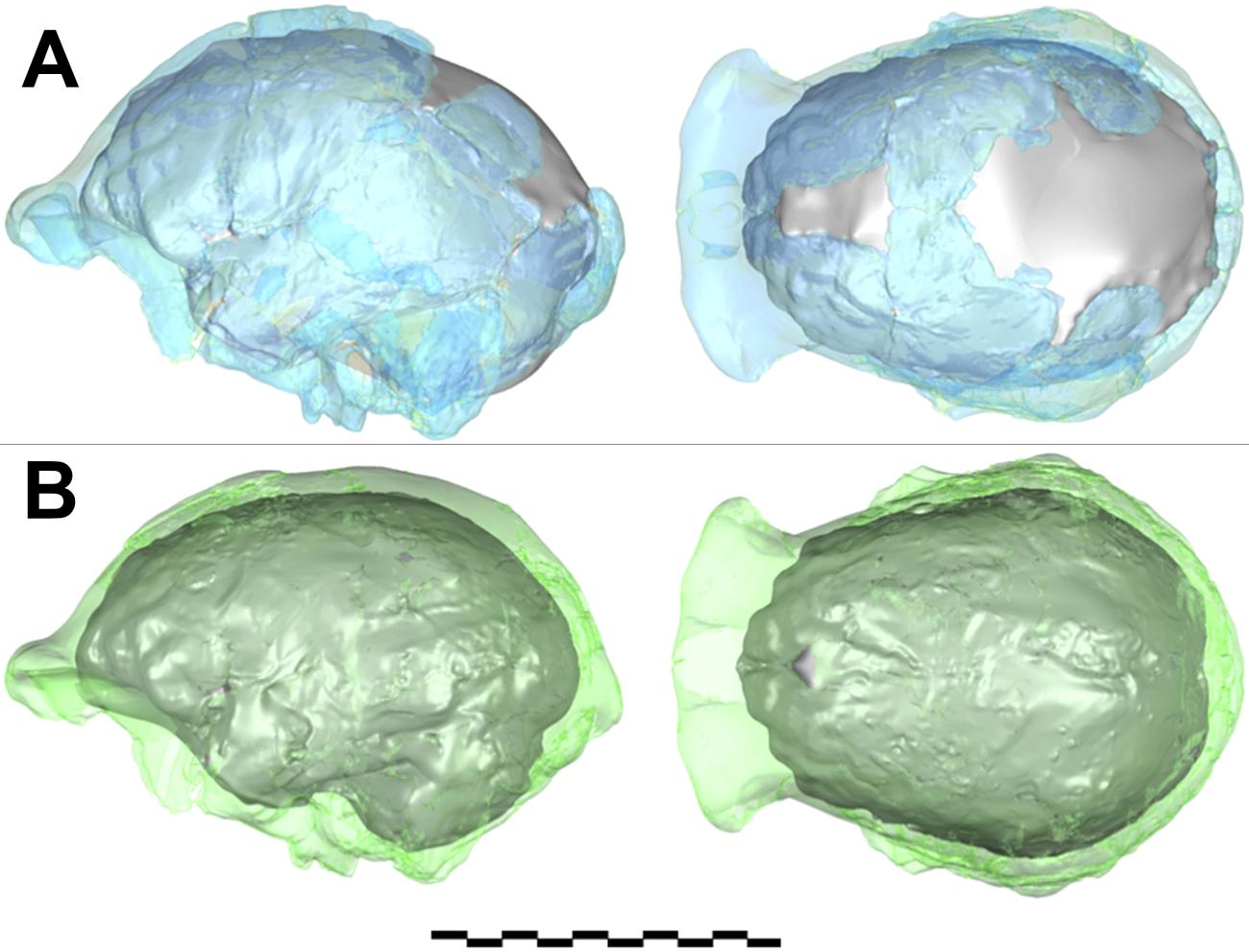
2) Figures. It is important to understand the way in which some quantitative information is obtained from fossils. For example, in Table 1 of the paper [7] the cranial capacity (a measure of brain size) of Homo naledi is indicated as 513cc. In the methods section of the paper a good explanation is offered of how this figure was obtained. The process involved merging two sets of cranial remains on two different 3D models, interpolating areas where no data were available, and then making an average between the two calculated volumes (Fig. 3). This process of calculation gives a fair indication of the average brain size that seems representative of at least 4 individuals, but it is not a number obtained from direct measurement of a complete specimen. Another example is the estimate of body mass. The paper estimated the body mass of Homo naledi from bones of eight individuals, obtaining a range between 39.7 kg and 55.8 kg [8]. These numbers were calculated by measuring the breadth of femur bones at standard locations and applying to the measurements a regression equation based on modern human samples, where the body mass and femoral breadth of human individuals can be accurately established and correlated. This is standard procedure, but it is clear that the estimated body mass will vary if different modern human samples are used as reference and depending on the specific femoral measurements used. For example, using one of the same measurements adopted in the paper [9] but with a regression equation based on a different human sample [10] a range between 33.09 kg and 40.05 kg is obtained, which is less than the body mass values presented in the paper. Moreover, the assumption that the correlation between femoral breadth and body mass in modern humans is the same as in anatomically different fossil hominids is obviously not testable and probably an approximation at best.
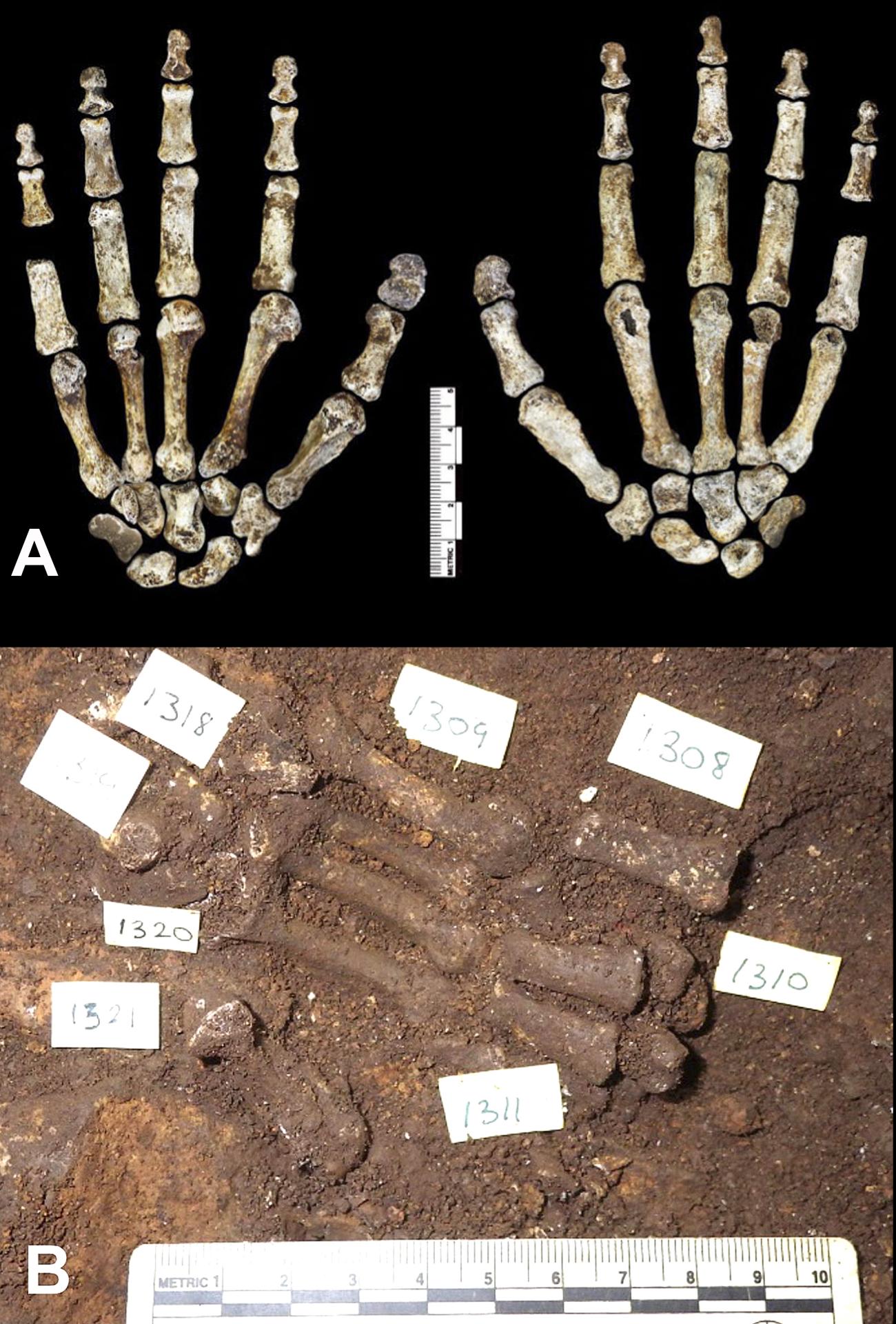
3) Attribution. Because most of the remains were not found in articulation (but see an exception in Fig. 4), it is difficult to establish which bones belong to the same individual. Paradoxically, if only one skeleton had been represented in the assemblage, even partial remains could have been used for direct measurement of actual body proportions. Having abundant disassociated material allows for a better understanding of the range of morphological and size variability in a population, but reconstructions of body proportions can be more complicated. Another possible complication with disassociated remains could be treating a mixture of two distinct types of hominid (e.g., one more human-like another more australopith-like) as if they belonged to one single type. The authors of the paper make a good case to exclude this possibility. They note that “in all cases where elements area repeated in the sample, they are morphologically homogeneous, with variation consistent with body size and sex differences within a single population” and that “distinctive morphological configurations,” including “traits not found in hominin species yet described,” are identical in the recovered specimens. Their conclusion is that “these considerations strongly indicate that this material represents a single species, and not a commingled assemblage” [11]. However, even if this reasoning is indeed compelling, the theoretical possibility of mixing of two types remains, and was indicated in commentaries of the discovery by renowned paleoanthropologists Jeffrey Schwartz (University of Pittsburgh) and Ian Tattersall (American Museum of Natural History) [12].
4) Behavior. A final demonstration of the gap between data and interpretations relates to the setting in which Homo naledi fossils were found. The remoteness of the cave chamber (Fig. 2) and the fact that only hominid remains were found there has resulted in the assertion that this is an example of intentional disposal of carcasses. However, this scenario is only a possibility and cannot be established with certainty. In fairness to the authors, their conclusions acknowledge that other scenarios are possible: “Both the mass mortality or death trap scenario […] and deliberate disposal hypothesis are considered plausible interpretations and require additional investigation,” although the authors do express a preference for the deliberate body disposal inference [13]. Unfortunately, the caution and balance expressed in the paper did not translate in the remarks made by the authors to the media, where biased statements like these were released: “We have just encountered another species that perhaps thought about its own mortality, and went to great risk and effort to dispose of its dead in a deep, remote, chamber" [14]; “We can tell that this wasn't a social group that died through some sort of catastrophe in the cave" [15]; “We have, after eliminating all of the probable, come to the conclusion that Homo naledi was utilising this chamber in a ritualised fashion to deliberately dispose of its dead” [16]. Claims of ritualistic behavior, thoughts about own mortality, and assertive exclusion of the mass mortality scenario of a social group not only represent a stretching of the available evidence but in the latter case are also in plain contradiction with the conclusions of the paper.
The Homo agenda
The word Homo carries high significance, because it is part of the scientific name of humans. If an extinct type is placed within the genus Homo, it means that it shares closer similarity to humans than to any other known group of organisms, including australopithecines. Therefore, the choice of a name is not just a trivial technicality but conveys a sense of how the discoverers of a fossil would like it to be perceived. For evolutionists, something called Homo automatically takes a certain place in the alleged evolutionary sequence from australopithecines to humans. For creationists, something called Homo automatically falls within the morphological variations of a certain original created type. Both groups would want to avoid calling Homo something that is not.
The discoverers of the Raising Star cave fossils had to address two questions when choosing how to name the remains: 1) do these fossils represent a new type? 2) If so, to what is it most similar? By choosing the name Homo naledi, they answered the first question in the affirmative, and suggested humans as the answer to the second question. Although this choice might be legitimate, it is important to understand that it represents a specific interpretation of data and that other interpretations may suggest different conclusions to both questions. On the subject of this being a different type, for example, paleoanthropologist Tim White (University of California, Berkeley) considers, at an early stage of assessment, that these remains could be included within the variability of the already established species Homo erectus [17].
On the choice of the attribution to the genus Homo, the authors base their decision on human-like anatomical aspects described from the lower limb, hand, and teeth, as well as on the general shape of the cranium [18]. However, emphasis is deliberately placed on these characters while many other skeletal traits (e.g., pelvic area, shoulder region, ribcage) are more australopith-like than human-like. Estimated brain size, for example, is not only small in absolute value when compared to modern humans but also its relative proportion to estimated body mass (a measure known as encephalization quotient) is about half that of modern humans and similar to that of australopithecines [19]. In some of the presentation of the results, it seems that the authors of the paper wanted to deliberately emphasize a link with humans rather than australopiths. For example, the paper states that “H. naledi has a range of body mass similar to small bodied human populations” [20], but an equally true statement would have been that the range of body mass for H. naledi is similar to body mass estimated for large australopiths [21]. The emphasis on intentional disposal of cadavers is also framed as a supportive argument to highlight behavioral complexity currently seen only in humans. In theory, attribution of fossil remains to a genus should be based on statistical treatment of different skeletal and dental measurements that unequivocally group specimens with distinct anatomical traits. In practice, the procedure is complicated by the lack of agreement on which skeletal traits should be considered diagnostic for Homo and by the fragmentary nature of the fossil record [22].
Human evolution: A linear progression or a tangled mess?
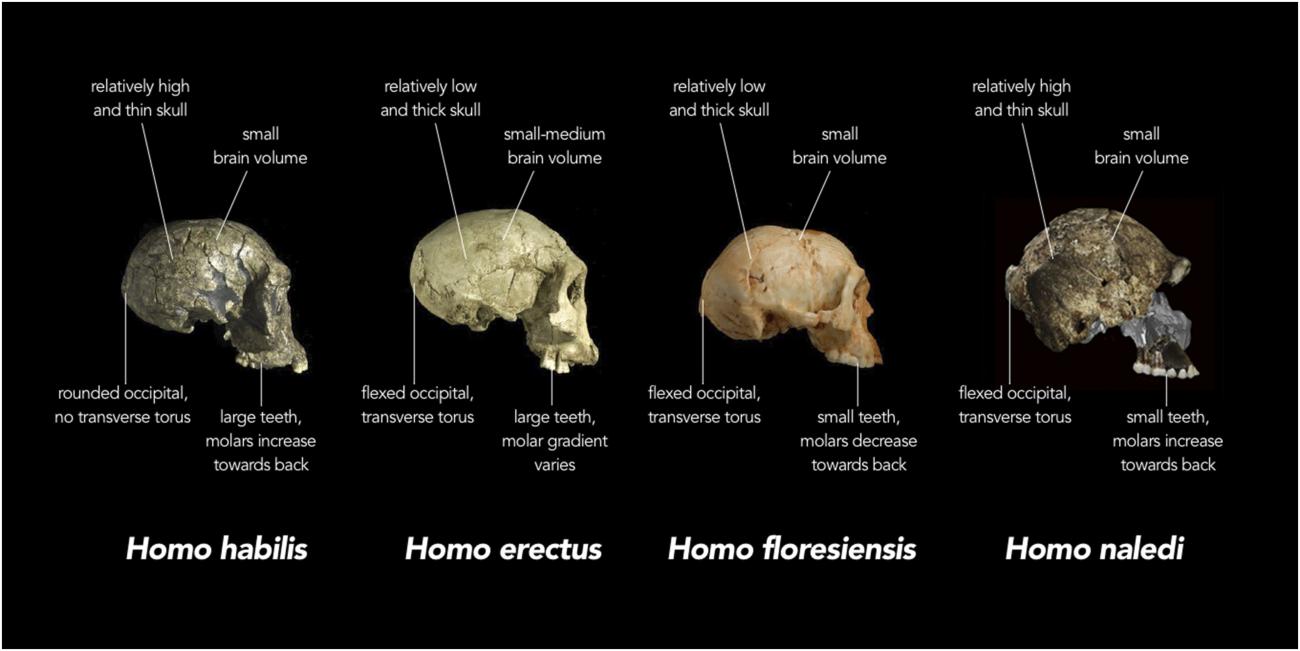
One of the most significant aspects emerging from the study of Homo naledi is its combination of anatomical features, some more similar to Australopithecus, others to Homo. This mosaic distribution of characters is seen at the level of the whole skeleton, with the trunk, shoulder, pelvis, and proximal femur being australopith-like and the lower limb and dentition being more human-like. However, mosaicism is also apparent in specific regions of the skeleton. For example, the hand shows an overall Homo-like morphology in the wrist bones but relatively long and markedly curved fingers (Fig. 4) . Teeth are small, like in humans, but with molar size proportions found in australopiths. Skull shape is more rounded, like in Homo, but cranial capacity and encephalization quotient are small, as for australopiths. A similar mosaic distribution of characters has been documented in other fossil hominids, such as Australopithecus sediba [23] and Homo floresiensis [24].
In addition to the mixture of skeletal traits observed at the level of an individual skeleton, recent discoveries from eastern Europe have revealed that a suprising amount of morphological variability was also present among different individuals within the same hominid population [25]. When considered together, these findings present a significant challenge to the classic scenario of sequential human evolution, popularized by the iconic procession of silhouettes gradually rising from ape-like to human stance. No simple, congruent, linear trajectory can be established for the development of individual human skeletal traits (Fig. 5) . On the contrary, human-like functional complexes appear with almost “random” distribution in conjunction with australopith-like characters in different organisms and among individuals in the same population. This paradigm shift is being acknowledged in important review papers. Antón et al., for example, maintain that “dynamic environments favored evolutionary experimentation and the coupling and uncoupling of biological variables, which governed against any simple transition from Australopithecus to Homo” [26]; they also talk of “intriguing shuffling of derived and plesiomorphic traits and biological variables that likely characterized the early evolution of Homo” [27], meaning a puzzling mixture of modern and primitive traits from an evolutionary perspective. Similarly, Schwarz & Tattersall, conclude that “in contrast to Mayr’s austere linearity, we may find that human evolution rivaled that of other mammals in its evolutionary experimentation and luxuriant diversity” [28]. Words such as “intriguing,” “experimentation,” “shuffling,” and “luxuriant diversity” are strong indicators of a new unexpected trend and an implicit acknowledgement of how little is really known with precision about certain aspects of the hominid fossil record.
What could be the origin of this variability and mosaic distribution of characters in fossil hominids? As is often the case in historical sciences, only suggestions can be made. However, Antón et al. hint at three possible processes: 1) morphological and developmental plasticity related to environmental conditions. For example, availability of food and low risk of mortality can extend growth and delay puberty in humans, resulting in larger body size and slower maturation; 2) hybridization between different groups, meaning the generation of offspring from individuals belonging to distinct types that can interbreed; and 3) vicariance, meaning diversification of groups as a result of isolation due to migration or geographical barriers [29]. It is interesting to realize that these same mechanisms would work very well also within a creationist framework, where australopith and human types would be expected to diversify in a post-flood dispersal.
There is a final important corollary of the pattern of mosaic distribution of traits that needs to be highlighted. As pointedly remarked by Berger et al., because of the non-predictable combinations of characters apparent in different fossil hominids “we must abandon the expectation that any small fragment on the anatomy can provide singular insight about the evolutionary relationship of fossil hominins” [30]. In other words, a reconstruction of alleged evolutionary relationships built on comparisons of just one element (e.g., the cusps of a tooth, the shape of a mandible, the prominence of a brow ridge) would lead to contradictory results if entire skeletons were available.
Lessons to learn
The publicity given to the H. naledi discovery will certainly raise questions on the subject of human origins. This great opportunity for deeper reflection on the subject will be beneficial if misconceptions can be avoided. Here are some important points that should be considered by and for those sharing a creationist perspective:
1) Is Homo naledi the “missing link” between australopithecines and humans? Irrespectively of what the media would like us to believe, the answer is no. First of all, the Homo naledi fossils have not been dated and their stratigraphic placement is unknown. They could very well be a relatively recent hominid group, therefore irrelevant in the alleged evolutionary emergence of Homo. Even if their stratigraphic position was determined to be between australopithecine and Homo fossils, H. naledi does not present a transitional arrangement of morphological traits, but, as we have seen, a mosaic combination, therefore not exemplifying a hypothetical pathway of gradual sequential changes expected in Darwinian evolution. A most significant gap still remains in the fossil record of hominids, making the transition from australopithecines to humans an interpretation of very scanty data.
2) What was H. naledi? A human or an australopith? As discussed earlier, this is a matter of interpretation. Probably, creationists will be inclined to see it as an australopith-type variant, especially in light of its small brain size, with adaptations for bipedalism and diet-driven modifications in dentition [31]. The reality is that we just don’t know. There are several viable options from a creationist perspective. H. naledi could be a hybrid between two types, a different type of primate, or an environmentally-driven variant of either the australopith-type or the human-type.
3) What was H. naledi not? A new discovery is also an excellent opportunity to remind what has NOT been discovered. H. naledi is not an anatomically modern human. So far, we have not discovered remains of modern-looking humans except toward the very top of the geologic column. H. naledi is not a giant human fossil. Unfortunately, hoaxes about giant human fossil discoveries continue to be popular among well-meaning but poorly informed Christians [32], and great skepticism should be exerted when presented with some sensational news on the topic. H. naledi is not likely to represent an antediluvian fossil. Its deposition within the surficial sediments of a cave places its existence at the very end of the sequence of geological events that formed the present landscape.
4) What larger lessons can be learnt from this amazing discovery? Certainly, the most instructive lesson is that every seeker of knowledge will always benefit from a clear understanding of the difference between data and interpretations. Biblical creationists need to be aware of the powerful philosophical presuppositions at work when presenting new discoveries, often rooted in an evolutionary worldview. This might lead to intentional or unintentional bias in the presentation of results or in the popularization of their implications. As we sharpen our critical eye, we should continue to work confidently towards the construction of models that are compatible with a biblical worldview, maintaining a humble attitude in areas of incomplete understanding.
Ronny Nalin, PhD
Geoscience Research Institute
For an update on the subject of Homo naledi including information from two papers published in May 2017, see our blog post linked here
Footnotes
[1] Berger, L.R., et al., Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife, 2015. 4.
[4] Dirks, P.H., et al., Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife, 2015. 4.
[5] McKenzie, D. and Wende, H., Homo naledi: New Species of Human Ancestor Discovered in South Africa. CNN, September 10, 2015, http://www.cnn.com/2015/09/10/africa/homo-naledi-human-relative-species/.
[9] AP subtrochanteric breadth, Table 3, p.18 of Berger et al.
[10] Grabowski, M., Hatala, K. G., Jungers, W. L., & Richmond, B. G., Body mass estimates of hominin fossils and the evolution of human body size. Journal of human evolution, 2015. 85, 75-93.
[12] As reported in Barras, C., New species of extinct human found in cave may rewrite history. New Scientist, September 10, 2015, https://www.newscientist.com/article/mg22730383-700-new-species-extinct-human-found-in-cave-may-rewrite-history/.
[14] Reported in McKenzie and Wende.
[15] Reported in Johnson, M.A., and Jackson, H., 'Mind Blown': Is Human Ancestor Discovery the Long-Sought Missing Link? NBC News, September 10, 2015, http://www.nbcnews.com/science/science-news/mind-blown-human-ancestor-discovery-long-sought-missing-link-n425406.
[16] Reported in Sample, I., Homo naledi: new species of ancient human discovered, claim scientists, The Guardian, September 10, 2015, http://www.theguardian.com/science/2015/sep/10/new-species-of-ancient-human-discovered-claim-scientists.
[17] Reported in Sample and in Hartley, R., Some bones to pick, Times Live, September 18, 2015, http://www.timeslive.co.za/thetimes/2015/09/18/Some-bones-to-pick.
[19] Using the body mass and brain size estimates of Berger et al. and data end equations in Ruff, C.B., Trinkaus, E. and Holliday, T.W. (Body mass and encephalization in Pleistocene Homo. Nature, 1997. 387, 173-176) the estimated encephalization quotient for H. naledi is about 2.4, whereas for modern humans it is about 5.3.
[21] For example, when the same equations and measurement types used to estimate body mass in Berger et al. are applied to AL 333-131 (a femur bone of Australopithecus afarensis) the value obtained is ~61kg, which exceeds all the body mass estimates for H. naledi. Using multivariate statistical analysis, Grabowski et al. calculate average male mass for A. afarensis as 49.5 kg, which also overlaps with the estimates for H. naledi.
[22] See, for example, Schwarz, J.H. and Tattersall, I., Defining the genus Homo. Science, 349/6251, pp.931-932.
[23] Berger, L. R., de Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H. and Kibii, J. M., Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science, 2010. 328/5975, pp.195-204.
[24] Brown, P., Sutikna, T., Morwood, M. J., Soejono, R. P., Saptomo, E. W. and Due, R. A., A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature,2004. 431/7012, pp.1055-1061.
[25] Lordkipanidze, D., de León, M. S. P., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A. and Zollikofer, C. P., A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 2013. 342/6156, 326-331.
[26] Antón, S. C., Potts, R., and Aiello, L. C., Evolution of early Homo: An integrated biological perspective. Science, 2014. 345/6192, p.1236828-10.
[28] Schwarz and Tattersall, p.932.
[31] This is, for example, the position taken by E. Mitchell on an Answers in Genesis blog on the H. naledi discovery: Is Homo naledi a New Species of Human Ancestor? September 12, 2015, https://answersingenesis.org/human-evolution/homo-naledi-new-species-human-ancestor/
[32] The latest fabrication to which I was recently pointed by a friend, who had some hopes of its credibility, can be found at http://worldnewsdailyreport.com/5-meter-tall-human...