The Intrigue of the Questions about the Bridgewater “Fossil Forest,” Victoria, Australia
Science Note by Clyde L. Webster, Jr. Geoscience Research Institute
WHAT THIS ARTICLE IS ABOUT
Fossils offer intrigue to everyone, from child to adult, and raise many questions. What is it? Where did it come from? How old is it? How did it become a fossil? and many more. Sometimes these questions have easy answers, and other times almost none of them can be answered. The Bridgewater "Fossil Forest" fossil structures fall into this latter category. Field evidence is examined in light of the current theories of the Bridgewater structures. As the data are examined almost none of the usual questions about fossils can be answered. Instead of answering questions, this study has only heightened the intrigue by offering more unanswered questions, such as their origin: transport (allochthonous) or in growth (autochthonous)?
Within the Pleistocene Bridgewater Formation on the western coast of Cape Bridgewater, Victoria, Australia, there are located several square kilometers of unique structures. Over the years these structures have been interpreted in many ways. [1][2][3] They are not only fascinating to the scientist, but also intrigue the layperson. Because of their uniqueness, protection and management of this area has come under the purview of the Department of Conservation and Natural Resources.
The basalt cliffs of Cape Bridgewater tower some 100 meters above the restless ocean. Overlying the black basalt is a reddish fossil soil horizon that has been exposed by erosion and consists of remnants of calcarenite cliffy benches. In scattered areas above these benches is exposed an additional somewhat younger red soil zone. Within these exposed fossil soils are structures resembling small upright trees, saplings, stumps and roots. The upright trees are found in the lower horizon, while above one finds recent branch-like fossil casts, some of which contain unmistakable woody material within their core. These recent uppermost fossils are distinct from the large tree-like structures below in that they are light beige in color, are directly associated with current local vegetation, and seem to be only a surface phenomenon.
One of the earliest references that mentions the fossil "forests" of the Bridgewater Formation is that recorded by Charles Darwin. This record was drafted during his famous trip on the H.M.S. Beagle:
One day I accompanied Captain Fitz Roy to Bald Head; the place mentioned by so many navigators, where some imagined that they saw corals, and others that they saw petrified trees, standing in the position in which they had grown. According to our view, the beds have been formed by the wind having heaped up fine sand, composed of minute rounded particles of shells and corals, during which process branches and roots of trees, together with many land-shells, became enclosed. The whole then became consolidated by the percolation of calcareous matter; and the cylindrical cavities left by the decaying of the wood, were thus also filled up with a hard pseudo-stalactitical stone. The weather is now wearing away the softer parts, and in consequence the hard casts of the roots and branches of the trees project above the surface, and, in a singularly deceptive manner, resemble the stumps of a dead thicket.
One of the most complete studies of the Bridgewater Formation and the fossil structures was prepared by N. Boutakoff in 1963. [4] However, Boutakoff devoted most of his efforts to the description of the geology of the Bridgewater Formation and the nearby Portland area, only briefly mentioning the fossil structures found within the Bridgewater Formation itself. In his description, Boutakoff does address the relationships of the tree-like structures and the possibility of these structures being solution pipes. As a result of his investigations he concluded that the popular name "Petrified Forest" is fully justified, even if the mechanics of formation cannot be fully described or understood.
Another excellent description of the geological setting of the Bridgewater Formation is found in the treatise "Geology of Victoria," edited by J.G. Douglas and J.A. Ferguson. [5] This treatise is devoted mostly to the geology of the area, and makes only a brief mention of the fossil tree-like structures found within the Bridgewater Formation. In this reference, the description and interpretations given by Boutakoff are mentioned and accepted without question. The main value of the two references mentioned above is their contributions to the understanding of the geology and geomorphology of the Portland area and the Bridgewater Formation.
A short passing reference to these fossilized structures can be found in the VULCON Guidebook of the Vulcon 1995 20th Biennial Conference. On p 96 these structures are simply referred to as "solution pipes." [6]
With the exception of three or four small fossil structures located in nearby road cuts, the major exposed fossil structures are found between 38º 21' and 38º 23' south latitude. These fossil structures seem to be bounded to the north by a white outcropped limestone very similar to the major limestone outcrop located at the east side of the Bridgewater Lakes. All the major fossil exposures are located west of the Cape Bridgewater Fault.
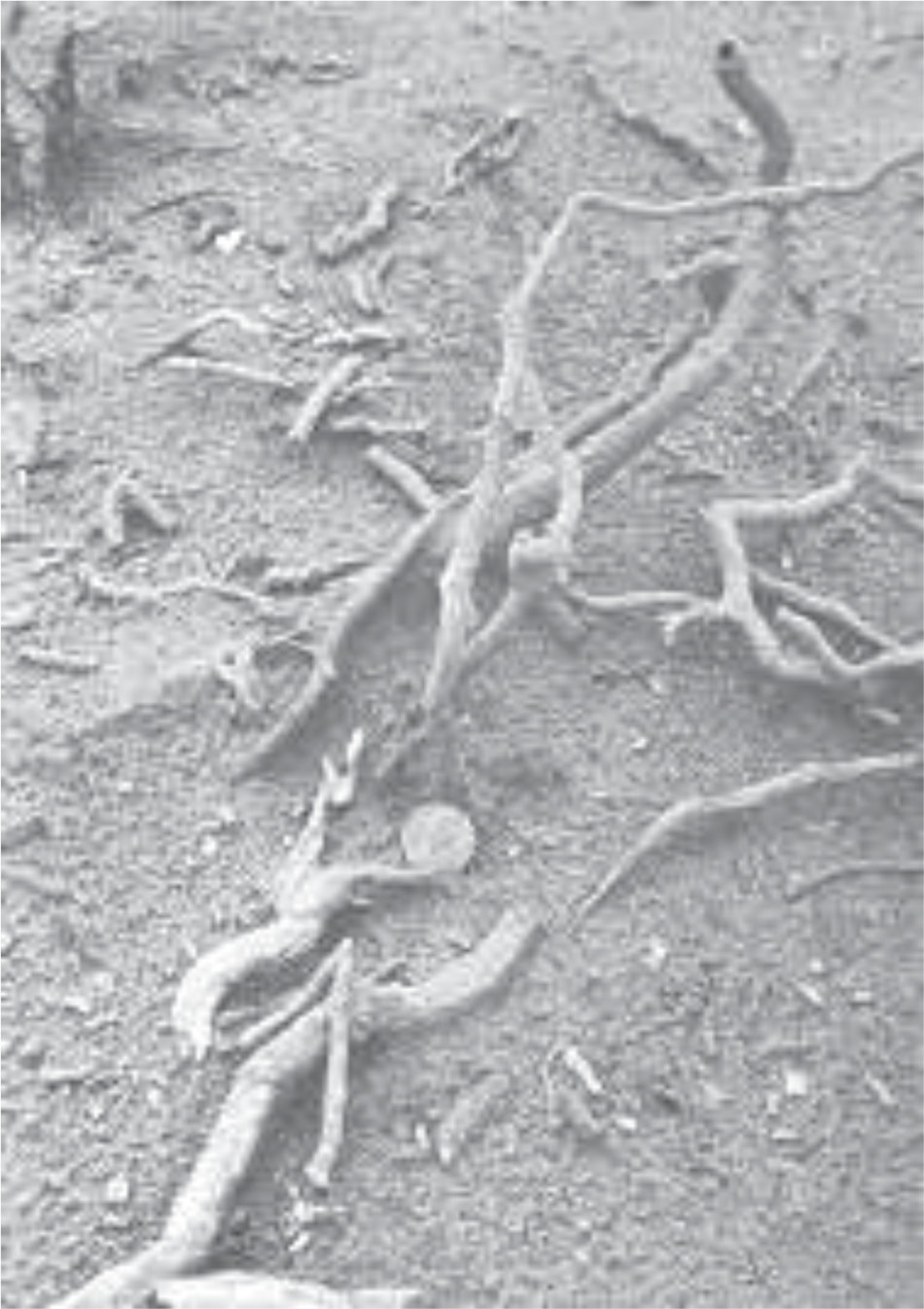
As mentioned earlier, there are two distinct types of fossil structures: large (100 to 500 mm) diameter molds, found in the lower layers (see Figure 1), and small (10 to 80 mm) limb and root casts, found higher up (see Figure 2). The large molds, when exposed, may attain heights of 1 to 2 meters and may contain a uniform sandy/fat clay (similar to a deeply weathered tropical soil in color but not texture) infill. The limb and root casts which still contain the fibrous matter normally do not exceed 1 meter in length.
Figure 1. Multiple fossil structures resembling small upright trees at Cape Bridgewater. Some of these structures may reach a height of 2 m and a diameter of 400 mm.
The large vertical molds are almost exclusively found in open barren terrain while the overlying modern limb and root casts are found in active dunes and bordering local ground cover. Informal observation immediately suggests that these limb and root casts are in near-growth density and position. On the other hand, such conclusions cannot be so easily drawn for the large molds.
Figure 2. Limb and root casts found within local active calcareous dunes at Cape Bridgewater. When broken open, many of these casts still contain fibrous material in the center.
Only large mold structures are found below the cliff tops along the Great South West Walk trail. Near the base of the cliffs, the bottom of these large mold structures can be seen abruptly truncating at the top of the basalt basement of Cape Bridgewater. Two or three layers of these large casts may be observed in several locations below the cliff tops. There is no communication between these multiple layers. No multiple layers of the overlying limb and root casts have been observed.
The sandy/fat clay infill of the large molds was removed from several of the molds at various locations within the main exposed area. There was no evidence of infill stratification or mixture with detrital debris in any of the molds. The infill matrix exhibited the same soft, non-compacted consistency regardless of the location.
When exposed, the lower ends of the molds are often truncated with a rounded slightly bulbous end (see Figure 3). Root systems, when present, can almost exclusively be traced to this bulbous end. The multiple layer molds also exhibited this type of truncation.
Figure 3. Bulbous truncation common to many of the large vertical structures at Cape Bridgewater.
In one specific area several molds with double-ring structures (a mold within a mold) were noted. The inner ring is approximately 30-40 mm smaller in diameter than the outer ring.
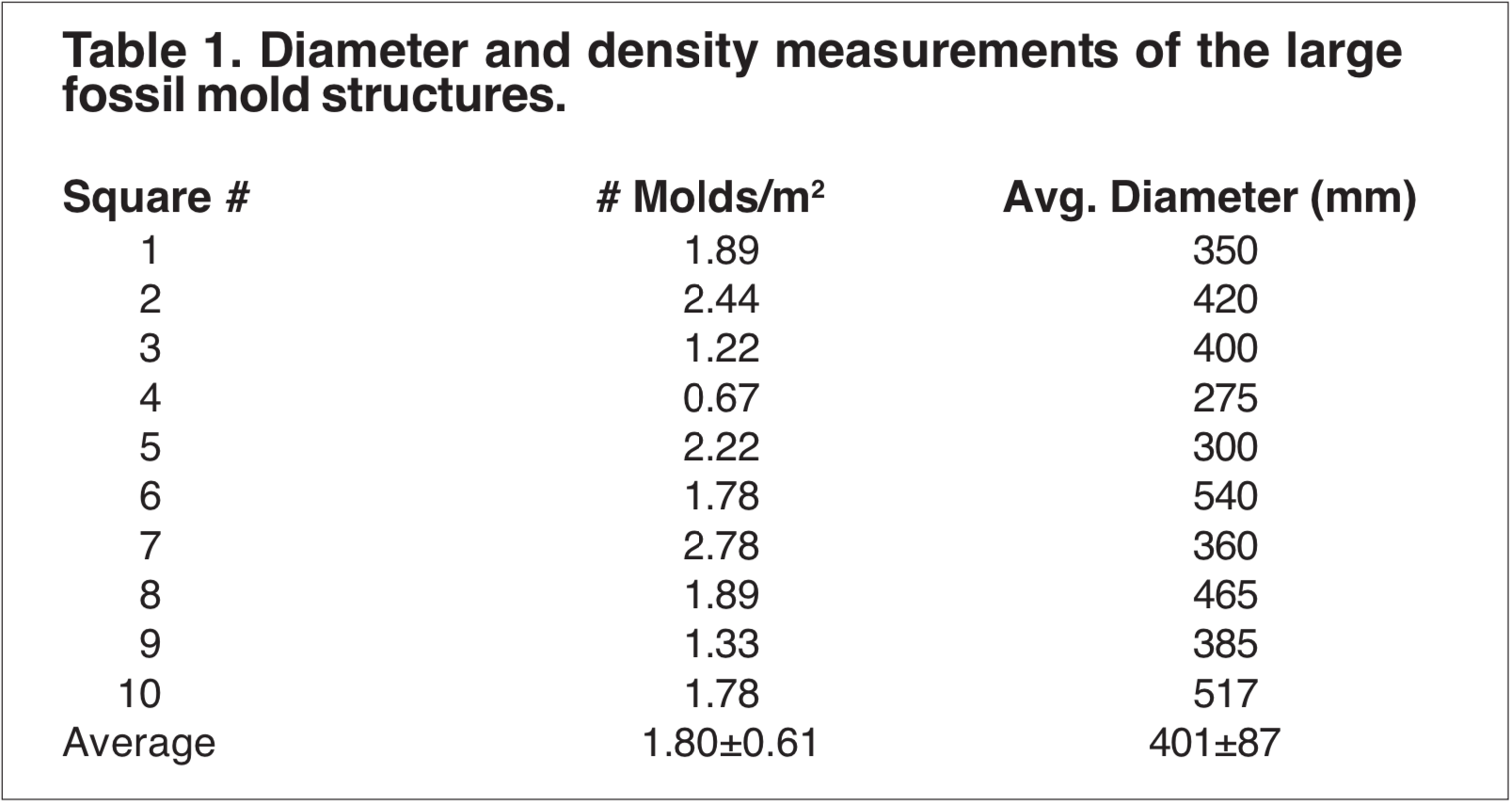
Growth density measurements for the large mold structures were obtained by randomly selecting 3×3 meter sites within the formation and obtaining a number count. The diameter of each mold was also measured and recorded (Table 1).
The data were initially examined to determine if they were consistent with either a fossilized autochthonous (in-growth position) forest hypothesis or a solution pipe hypothesis.
The solution pipe hypothesis seems to be the easiest hypothesis to evaluate. If these structures are solution pipes the following questions need to be answered:
How can one account for such a dense concentration of pipes, over such a large area?
If these structures are pipes, why are the pipes so close together without showing evidence of collapse or coalescence with one another?
How can multiple layers of pipes be accounted for, especially when there is no evidence of communication between layers?
If these structures are pipes, how can bulbous terminations be accounted for, especially those structures which seem to have roots radiating from the terminations?
In addition to the uniform termination of the pipes, how can we account for the fact that many of these structures remained hollow? When the solutions ceased to flow, why didn't the pipes just fill up and harden?
In attempting to answer the above questions it becomes obvious that these features require an explanation other than a solution pipe hypothesis. The most popular hypothesis is an autochthonous (in growth position) fossilized forest. This hypothesis seems to be supported by the general size, orientation, and shape of the large mold structures. The presence of roots extending from the base of some of the molds into a presumed paleosol would also tend to support the tree hypothesis. However, there are a number of other questions about this hypothesis which need to be addressed:
If these structures represent normal trees, why are the vast majority of molds hollow?
How can one account for the close spacing density if these structures represent a normal autochthonous forest?
If these structures represent an autochthonous forest, how can one account for the multiple layering of these structures?
Some of these questions were also raised by Boutakoff in 1963. [7] It would seem that none of the standard hypotheses adequately address all of the data. This is especially true when the small limb and root structures are included with the large mold structures.
In order to accommodate all or nearly all of the field observations and data, this investigator is proposing a two-part model consisting of both an autochthonous and an allochthonous (transported) interpretation of the data.
The autochthonous deposit is recent. It includes the upper beige calcareous limb and root casts. Excavation of the limb casts reveals the root casts that are in direct communication with the limb casts and often contain fibrous woody matter. These beige structures have not been found within the red paleosols of the larger vertical tree-like structures and appear to be derived directly from the calcareous dune sands. The limb and root casts border similar existing ground cover and conform to local growth patterns and density. These beige cast structures are also approximately the size and shape one would expect from the fossilization of the current ground cover.
A plausible model for the formation of the limb and root casts has been outlined by Boutakoff. [8] He suggests that during initial formation, the limb or root is simply enclosed in calcareous sand, which results in an increase of the original diameter. Secondly, the enclosed matter decays, either partially or totally, and becomes calcified. The resulting calcified structures are what is observed in the field today.
Since the surrounding cast material is similar in composition to the local migrating dunes, the following scenario can be proposed:
The migrating dune encroaches living ground cover, eventually inundating and killing the plants.
As the plant material decays, the organic acids exude into the surrounding sand and form a lithic calcareous sand cast about the limb or root.
After formation of the cast, wind erosion exposes the casts.
The thickness of the cast is controlled by the amount of organic acids produced during decomposition. In other words, the size of the cast is proportional to the initial size of the root or limb and the degree of decomposition.
The lower, large mold, allochthonous deposit of the model comprises the largest portion of the Bridgewater Fossil Formation and is the most difficult to understand. The allochthonous segment of the model contains only large mold casts. Evidence supporting the allochthonous position comes from the fact that there are multiple, successive layers of these structures without any evidence suggesting a fossil soil zone. Additionally, in order to accommodate the growth canopy, the growth-density for an in-situ forest should be much less than the measured mold structure density, unless the flora was similar to the horsetails. Furthermore, an allochthonous model for the large casts would be more compatible for structure emplacement in limestone than the autochthonous model because the limestone could also be a product of the transporting environment.
Before an acceptable depositional scenario can be proposed, the following questions need to be answered:
If the mold structures were in-situ, why aren't more root systems found?
Why are a large percentage of the mold structures hollow?
Where did the non-stratified infill matrix originate?
What type of flora is represented by the mold structures?
What is the source of the surrounding limestone?
If not allochthonous, what was the mechanism of deposition?
When all of the field data are taken into consideration it seems logical to divide these fossil structures into a modern segment and a prehistoric segment. The reasoning behind this division is two fold. First is the distinct difference in the calcilithic agents. The modern segment seems to be composed of material similar to the calcareous dunes in the locality. The prehistoric large mold structures are composed of a totally different calcilithic agent, as determined from composition and color. Second, the root and limb casts are similar to the local living flora, and seem to be directly associated with such flora. In other words, the root and limb casts seem to be a surface phenomenon only. In contrast, the large mold structures seem to have no modern-day analogue. Because of their superposition, all of the fossil structures (modern and prehistoric) had to occur after the deposition of the Iddingsite basalt flows that form Cape Bridgewater and Cape Duquesne.
Interpretation of the multiple layering of the large mold structures, coupled with the bulbous terminations and lack of roots or rhizome structures for these molds, would lend itself more readily to a transport mechanism. Transport and deposition could be directly or indirectly associated with the subsidence of the Nelson caldera, as proposed by Boutakoff. [9] However, the hollow features of these molds is problematic to a transport mechanism unless the structure of the flora itself is composed of similar-length segmented tubes.
An enigmatic feature of the prehistoric large molds is the red sandy/ fat clay infill. What is the origin of this material? Why is there no evidence of stratification of this infill material within the mold structures? Why is there a distinct lack of detrital matter within this infill matrix? One of the few logical explanations for this infill material is to suggest that this material originated from a single event and a single source, and that the filling process was rapid. Because the infill is not lithified and exhibits a different mineral composition than the surrounding shell, the major fossilization mold probably occurred before infill.
Are there any flora analogues for the mold structures found in other Pleistocene deposits in the surrounding area? Before this question can be answered, one needs to know if there is any possible prehistoric flora analogues similar to these mold structures, irrespective of their position within the geologic column?
A modern day analogue can be found in the horsetails. Horsetails are in the family Equisetaceae, which are found within the geologic column from the Permian to the recent. [10] Horsetails have hollow segments, closed at the upper and lower ends. The lowest of these segments has a bulbous termination from which the root mass extends. It should be noted that the average diameter of modern horsetails in the western hemisphere is on the order of 10 mm rather than 400 mm. However, ancient horsetails of flora do reach the size of these large molds. [11] The similarities of modern horsetail and the large mold structures are impressive. A study conducted on flotation characteristics of modern horsetail by H. G. Coffin [12] suggests that upright position and segment separation of Equisetum fossils may be associated with transport and not restricted to in-growth position. This is especially true if there is an absence of basal rhizome structures. In several sites, the large molds exhibit structural features that may be interpreted as segmentation. Segmentation can also be implied from the fact that the large mold structures, where completely exposed, seem to have equal lengths.
A review of J.G. Douglas' book "What Fossil Plant Is That" [13] and "Geology of Victoria" [14] suggests that members of the Equisetaceae family are not found above the Cretaceous in Victoria. This may not be a problem with a catastrophic transport mechanism. It may be that the Equisetaceae grew in the inland Malanganee swamps, and were simply stripped off and deposited at Cape Bridgewater. Before such a mechanism is acceptable, more research needs to be conducted in the Portland, Victoria area.
CONCLUSIONS
As one reviews the field observation data and the geological setting of the Cape Bridgewater area, a catastrophic transport/deposition hypothesis seems to currently offer a better explanation for the large molds than does an in-situ growth hypothesis. A transport/deposition hypothesis could account for the multiple layers of the large mold structures as well as the unique infill matrix. Additionally, a transport/deposition hypothesis would be more compatible with these large mold structures being found in the limestone cliffs directly above the Bridgewater basalt.
The beige root and limb casts are a recent phenomenon directly associated with the calcareous dune migration. The source material for these casts seems to be the local flora rather than material transported in from a different area. These modern fossils do not require any unusual explanation such as transport or deposition.
The value of this study extends beyond the specific locality in that it supplies additional information in understanding the paleoecology and sedimentation history of the Pleistocene epoch of the southern hemisphere.
ENDNOTES
[1]Darwin C. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle around the world. London.
[2]Woods JET. 1862. Geological observations in South Australia. London: Longman, Green & Co., p 171.
[3]Fairbridge RW. 1954. Quaternary eustatic data for Western Australia and adjacent states. Proceedings of the Pan Indian Ocean Science Congress, Perth, August 1954, Section 4, p 64-84. Further references will also be found here.
[4]Boutakoff N. 1963. The geology and geomorphology of the Portland area. Memoirs of the Geological Survey of Victoria 22:52-58.
[5]Douglas JG, Ferguson JA, editors. 1988. Geology of Victoria. Victorian Division, Geological Society of Australia, p 365-373.
[6]Braddeley G, editor. 1995. VULCON guidebook: lava features and limestone karst of Victoria and South-Eastern South Australia. 20th Biennial Conference, Australian Speological Federation, Inc., Hamilton, Victoria, 02-06 January, 1995.
[10]Stewart WN, Rothwell GW. 1993. Paleobotany and the evolution of plants. 2nd ed. NY: Cambridge University Press, p 198-212, 511.
[11]Meyen SV. 1987. Fundamentals of paleobotany. NY: Chapman and Hall, p 89.
[12]Coffin HG. 1971. Vertical flotation of horsetail (Equisetum): geological implications. Geological Society of America Bulletin 82:2019-2022.
[13]Douglas JG. 1983. What fossil plant is that? A guide to the ancient floras of Victoria. Field Naturalists Club of Victoria. West Melbourne: Jenkin Buxton Printers.
FRONT COVER: Enigmatic fossil structures found in the Pleistocene Bridgewater Formation of Cape Bridgewater, Victoria, Australia. BACK COVER: Surface view of one of these structures that displays annular layers. For further discussion about the origin of these intriguing structures see the General Science Note by Clyde L. Webster, Jr., beginning on p 50. Photographs by the author.
FRONT COVER: Enigmatic fossil structures found in the Pleistocene Bridgewater Formation of Cape Bridgewater, Victoria, Australia. BACK COVER: Surface view of one of these structures that displays annular layers. For further discussion about the origin of these intriguing structures see the General Science Note by Clyde L. Webster, Jr., beginning on p 50. Photographs by the author.