FORAMINIFERS IN THE FOSSIL RECORD: IMPLICATIONS FOR AN ECOLOGICAL ZONATION MODEL
by Tammy Tosk M.A. in Geology San Bernardino, California
WHAT THIS ARTICLE IS ABOUT
Foraminifers — shelled protozoans — are found fossilized in marine sedimentary rocks through most of the geologic column. They exemplify many of the problems of fitting the fossil record into a short chronology. These problems include the existence of large numbers of specimens and thousands of named species, and apparent evolutionary sequences of simple to more complex forms. Any short-time creationist model must be able to explain these features.
Foraminifers are able to reproduce rapidly and are quite variable morphologically, depending on environmental conditions. These factors make it seem plausible that the numbers of species and individuals in the geologic column could have lived in the time since creation week. In addition, the general distribution of foraminifers in the fossil record resembles their depth profile in the ocean today, with benthic forms distributed through a greater variety of environments than planktonic species. Although not a comprehensive study, the results suggest that the distribution of foraminifers in the geologic column may be consistent with an explanation based on burial by a worldwide flood.
Do the sequence and diversity of foraminifers in the fossil record represent evolutionary development, or are there alternative explanations?
The fossils in the geologic column have been thought to be a record, although certainly not a complete record, of the development of life on Earth. Because the prevailing paradigm assumes that the current processes operated at current rates in the past, the fossil record is assumed to have been formed slowly as evolving plants and animals lived, died and were buried, as we observe today.
Scientists working under that paradigm look for different kinds of information than do those who assume that the fossil record is the result of a major catastrophe. Because they think that fossils buried low in the geologic column must be much older than and ancestral to those buried in the upper parts of the column, they look for similarities and differences indicating evolutionary relationships.
Many scientists also assume that the fossils lived in the area where they were buried and fossilized, only being transported before burial in ways similar to those observed today. Fossils would thus give information about the environment of the area where they lived and died.
If a major catastrophe such as the Noachian flood was involved, the fossil distribution would be the result of factors other than just time and evolutionary change. A scientist looking at the fossil record under a flood paradigm would assume that most of the fossilized plants and animals had been living contemporaneously, and this scientist would look for characteristics of the fossils that would explain their order of burial during a major catastrophe. Some of the information needed for such an interpretation is often included in the reports generally published, but much is not.
Species descriptions give information about the shape and structure of the fossil, but may not give differences in size, thickness and weight that would be significant in studies of their buoyancy and other transport characteristics. Differences in preservation, which could be indicative of extensive transportation, are generally only mentioned as problems for their identification. Stratigraphic occurrences (the vertical range of the fossil in the geologic column) and geographic occurrences (the locations where the species has been found) are generally given when known. However, only a small fraction of the sedimentary rock in the crust of the earth has been examined for fossils, so their true distribution and abundance can only be estimated.
Another problem with using published descriptions and stratigraphic data is that fossils are often placed in different taxa, even in different superfamilies, if they are found at different levels, even though they might be placed in the same genus or species if found together. It is therefore difficult to recognize potentially equivalent species in the geologic column.
Several questions must, however, be asked of the fossil record to determine if its formation could have taken place within a short period of time. To show the plausibility of a model in which a significant part of the geologic column was deposited during a one-year, world-wide flood and its aftermath, one must show that all the fossils in flood deposits could have been deposited or reworked during the flood events into the observed biostratigraphy, and that similar organisms living after the flood would have a biogeographic distribution and genetic variability consistent with the loss of most of their population during the flood events.
To fully answer all these questions for all the fossil groups would require many lifetimes of research. This paper will explore only the group with which I am most familiar the microfossils called foraminifers.
Significance of Foraminifers
Protozoans of the Order Foraminiferida have been used extensively for relative dating of marine sedimentary rocks. They are small, generally less than a millimeter in length, and often found in such abundance that hundreds of specimens can be recovered from a mudstone sample with only a few hours of work. Because they are easily recovered from drilling chips, they are used to correlate strata in oil wells. An extensive literature of taxonomy and stratigraphic occurrences has therefore been developed for economic as well as academic reasons.
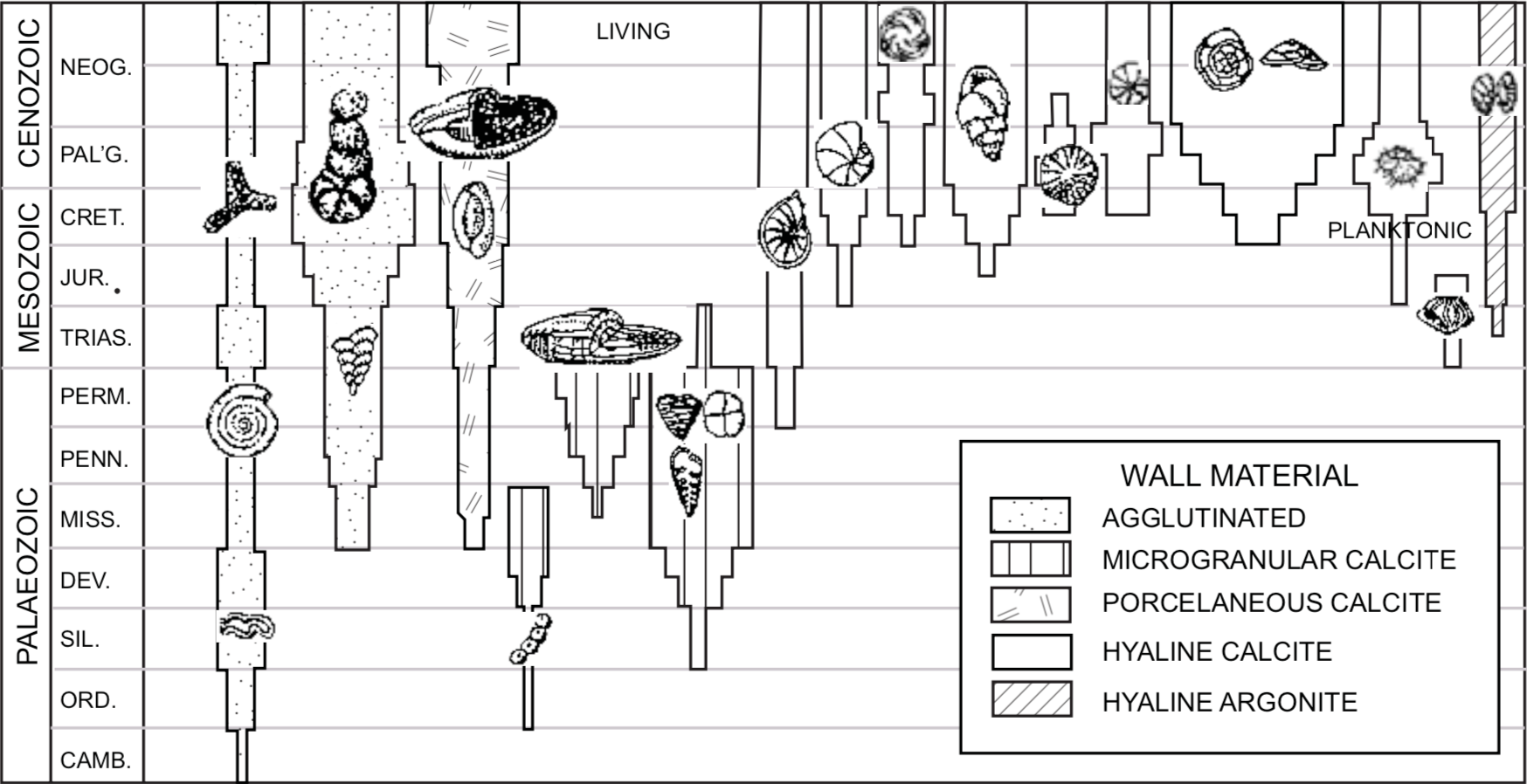
Foraminifers have more architectural diversity than any other fossil group. Many forms are long-ranging, being found through major segments of the fossil record. Other more specialized forms have very restricted ranges, and so are useful as index fossils. Foraminifers can therefore be used to correlate most marine sedimentary deposits.
Foraminiferal Architecture and Mineralogy
Foraminifer shells, called tests, have many designs including simple tubes, straight series of chambers, coils of chambers and even complex labyrinths. Their walls can be formed of foreign particles agglutinated in organic or calcareous cement, or totally of calcareous material secreted by the foraminifer. They interact with their environment through pores in the test wall and apertures of varying shapes and sizes, including some produced on long delicate necks. Foraminiferal taxonomy is based first on the wall mineralogy and microstructure, then on chamber arrangement, apertural shape and position, and ornamentation (Loeblich and Tappan 1964).
Foraminiferal Biology and Ecology
Foraminifers are abundant today and live in environments ranging from deep sea trenches to estuaries and even freshwater lakes. Planktonic and pelagic species live in the water masses at various depths, while benthic species live near, on or in the sediment on the sea floor. Some are permanently attached to the substrate or to the shell of another animal, but most are free living.
Pseudopodia, hair-like extensions of the foraminifer cell protoplasm, are used for locomotion, for gathering food and building materials, and, with the aid of adhesive material secreted by the foraminifer, for attaching to the substrate. In planktonic species the pseudopodia are symmetrically distributed around the test, which is also surrounded by frothy ectoplasm to aid in flotation. Because they have little control of their movement, planktonic species are passive feeders, randomly attaching to organic particles, algae, bacteria, or copepods which come into range of their pseudopodia (Loeblich and Tappan 1964).
Some foraminifers attach to the sea floor and agglutinate sponge spicules or other material into a tall branching structure to form a base for their network of pseudopodia (Haynes 1981). They are also passive feeders.
Free-living species of both infaunal habit (living within the sediment) and epifaunal habit (living on the sediment surface) are generally active feeders, searching for food particles and prey. In burrowing species, the pseudopodia may form a bundle at the aperture to move aside sediment grains. Foraminifers have been clocked at velocities of several millimeters per hour while moving through coarse sand (Haynes 1981).
Foraminifers are tolerant of extreme conditions, with forms living in areas of low oxygen levels, hyper- and hypo-salinity, and extremes of pH and temperature. In deep ocean environments where calcareous material is dissolved, foraminifers with agglutinated tests predominate (Haynes 1981). In nearly anoxic environments (<0.1 ml O2/l water), foraminifers may be flattened to increase the surface area through which to absorb oxygen (Douglas 1979). Their walls may be thinner and more porous, with less ornamentation.
Within a single species the foraminifers may have thick ornamented walls under normal oxygen concentrations, and thin, less-ornamented walls under low oxygen conditions. In a study of Pliocene foraminifers (probably post-flood), Hendrix (1958) broke open rocks containing both non-laminated massive and thinly laminated mudstone and examined the foraminifers exposed on the fresh surfaces. He found that the foraminifers in the massive sediments had thicker walls with more ornamentation, such as longitudinal ribs and keels, than specimens of the same species from the laminated sediments. Although many factors could influence the formation and preservation of laminae, they are often interpreted to indicate an environment with low oxygen levels (<0.3 ml/l water; Bernhard 1986).
If the changes are a response to the change in environment and do not permanently alter the genetic make-up of the species, the generations after the environment returns to normal will have their normal characteristics. Temporary changes in the morphology of a species resulting from changes in its environment are called ecophenotypic variation (Kennett 1976).
Foraminiferal Variability
Because of the many examples of variation in living and fossil forms, foraminifers are considered to be extraordinarily plastic (Kennett 1976). A foraminifer may contain enough genetic information to express many different forms, depending on the conditions. The effect on gene expression (shape and function) of environmental factors such as oxygen levels is a subject requiring further research.
Many of the so-called species in the fossil record were probably not separate biological species. A species is defined as a potentially interbreeding group. Fossil species can only be defined based on the characteristics of the preserved remains.
Species may be defined because of their stratigraphic utility. If a group of fossil foraminifers contains a continuum of morphologic forms and one form is found consistently lower in the geologic column than the other, separating them into two or more species would increase the precision of the biostratigraphy based on their ranges.
It is quite possible, therefore, for a few types of created foraminifers to have developed different forms to fill the various ecologic niches in the pre-flood seas without the necessity of genetic changes requiring long periods of time.
PLAUSIBILITY OF A FLOOD MODEL
Number of Fossil Foraminifers
The first question which must be asked of the fossil record to determine if it could have been formed in a flood scenario is quite simple. Are there too many fossilized foraminifers for them all to have lived and died within the short time allowed?
Answering this question quantitatively would require a detailed analysis of the entire geologic column to estimate the number of foraminifers preserved. Although foraminifers are found throughout the geologic column, they are actually quite sparse in most sedimentary rocks. In Paleozoic and early Mesozoic strata, most specimens are recovered from small fossiliferous areas. A micropaleontologist studying the Triassic, for example, may process samples from a dozen localities before finding any specimens.
The number of foraminifers that could have lived between creation and the flood is also difficult to estimate. The reproductive capability of foraminifers is among the highest on Earth, with a doubling time of 3.65 days (Berger 1976). The pre-flood conditions were likely good for rapid growth and reproduction. During and after the flood the turbid waters would have included high levels of organic matter and other nutrients needed to fuel growth.
Number of Species
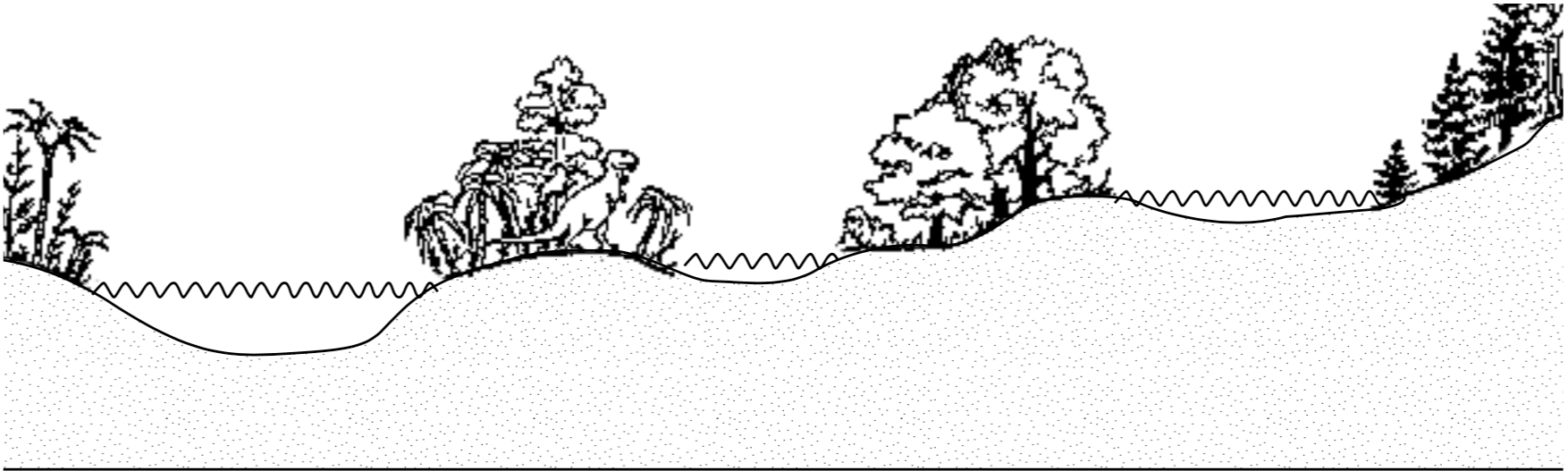
The oceans today contain species which are zoned by depth and other factors. If the pre-flood world contained several seas at different levels, as was proposed by Clark (1946), separate morphological species could have developed to fill the niches in those seas (Figure 1). Intensive differentiation could also have occurred during and after the flood events as conditions changed. With the inherent plasticity in foraminifer species discussed above, the many morphological species found in the fossil record could have developed in thousands rather than millions of years.
FIGURE 1. A representation of pre-flood environments as proposed by Clark (1946). The gradual erosion/burial of these ecologic zones by flood waters is used to explain the fossil sequence in sedimentary deposits. Representation not to scale.
Distribution in the Fossil Record
The stratigraphic distributions of foraminiferal taxa are fairly well described in the literature, but determining if that distribution could be the result of a catastrophic flood is another subject which would require years of research.
A significant problem arises because similar forms are classified differently if they occur at different stratigraphic levels. These cases are explained as iterative evolution, that is, the same form evolved repeatedly through geologic history. Thus classification is subjectively influenced by evolutionary theory. Repeated occurrences could be explained as easily by a catastrophic flood model. If the foraminifers found fossilized at various levels in the geologic column were living at the same time in different ecologic zones, species common to several ecologic zones would be found at several levels. Gaps in the record only indicate that the species was not present in the source area or the ecologic zone being buried at that time, not that it was totally extinct. No coincidence of repeated extinction and identical evolution is required.
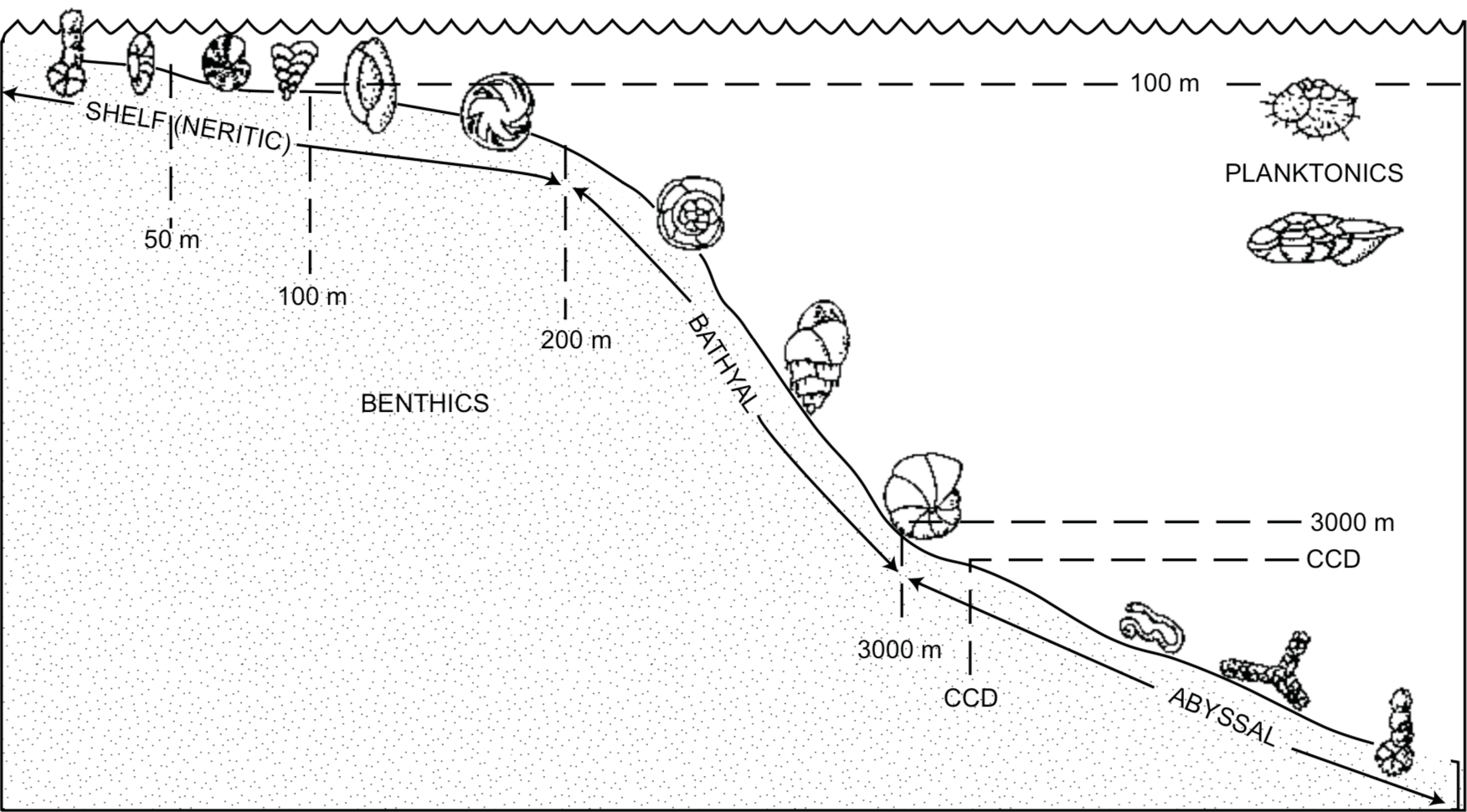
Ecologic zonation as developed by Clark (1946) would mean that foraminifers living in the lower seas or deeper parts of the ocean would be buried first as the sediments were redeposited by the gradually rising flood waters, while those from higher ecologic zones would be buried later The fossil record seems generally consistent with this model. Figures 2 and 3 show the distribution of foraminifers today and of fossils in the geologic column. Simple agglutinated forms that now live in environments ranging from the deep sea to estuaries, are found fossilized in Early Paleozoic and younger strata. Calcareous benthic species now predominate both in bathyal environments (Figure 2) and in Mesozoic strata of the past, and presently floating planktonic forms from a higher ecologic zone are abundant in the higher Cenozoic strata of the past.
FIGURE 2. Representative foraminiferal types in the ocean today. Depth data from Brasier (1980) for planktonic and nearshore environments, Bandy (1953) for bathyal depths and Schroder et al. (1988) for abyssal depths.FIGURE 3. Stratigraphic distribution of foraminiferal groups. Width of bar represents number of families. (Modified from Brasier 1980).
In the oceans today, calcareous material is dissolved below the carbonate compensation depth (CCD) pusually at a depth of about 4000 m, depending on carbon dioxide concentration. Neither benthic nor planktonic calcareous foraminifers are generally found below that depth on the abyssal plains or in deep sea trenches, because their calcareous shells would be dissolved. Agglutinated forms are dominant (Figure 2).
Agglutinated species are common in the Lower Paleozoic, and the benthic calcareous foraminifers found generally have thicker walls than forms higher in the geologic column. They could have lived near the pre-flood CCD where most calcareous forms, especially thinner-shelled planktonic species, would have been completely dissolved. Lower Paleozoic foraminifers are consistent, therefore, with the distribution expected by a catastrophic flood.
The fusulinids in the Upper Paleozoic, however, are an anomaly. Some species of fusulinids grew to volumes of more than 100 m3 (Ross 1979). Foraminifers which grew that large today have symbiotic photosynthetic algae living in their tests, and so must live within tens of meters of the ocean surface where sunlight is available. Large foraminifers from other groups live in shallow water tropical environments today; therefore, the fusulinids are interpreted also to have lived in a similar environment (Ross 1979), yet we do not find them at the top of the geologic column. Possibly they grew at the surface of pre-flood bodies of water of low altitude (Figure 1).
Planktonic foraminifers are not found in Paleozoic or Lower Mesozoic deposits. Even though living planktonic foraminifers float and would not be expected to be found in the early flood deposits, tests of those which had died before the flood should have been on the sea floor and should have been buried with those living there. Either they were not present in those ecologic zones, or they were not preserved as fossils. Because they have thinner, more porous tests than benthic forms, they could easily have been dissolved preferentially on the sea floor before the onset of catastrophic flooding, if their shells sank below the CCD.
Benthic hyaline calcareous foraminifers become abundant in the Mesozoic. Triassic and Jurassic foraminifers are generally not as well preserved as later forms. In Cretaceous strata, both benthic and planktonic forms are diverse and abundant, making it correlative with the upper bathyal zone of the ocean today.
Foraminifers older than the Cretaceous are generally widely distributed. A Triassic species may be found in both Australia and Idaho, but nowhere in between (Tosk and Andersson 1988). Cretaceous and younger foraminifers have distribution patterns correlative with modern assemblages (Sliter 1972). Under the prevailing paradigm, this would mean that the pre-Cretaceous seas were more cosmopolitan because modern hydrographic patterns and ecologic distributions had not yet developed. Continental fragmentation and sea-floor spreading during the Cretaceous are used to account for the development of modern oceanic patterns at that time.
In a flood model, however, this pattern is what would be expected. During the more violent stages of the flood events, foraminifers from a small area would be scattered widely over the earth. As the violence of the flood died down, foraminifers would not be transported as far and might even begin developing their own ecologic distribution patterns. Major deposition during and after the Cretaceous could have become localized in basins and at continental margins. Life for foraminifers may have returned to normal in less affected areas.
Post-Flood Foraminifers
Some foraminifers must have lived through the flood. Those that could survive such a catastrophe would be forms tolerant of turbidity and strong wave action, and possibly the juveniles of more delicate forms.
All major groups of foraminifers in the fossil record are represented by living forms except the complex fusulinid group which dominated the Paleozoic. As discussed above, they are interpreted to have lived in a tropical-type environment with low sediment output, and they may have had little tolerance for storm conditions. The agglutinated forms found in Paleozoic strata are found in many extreme environments today. They may have been better able to withstand the flood events.
No particular biogeographic distribution would be expected of foraminifers, unlike the animals which were saved in the ark and would have dispersed afterwards. Foraminifers would have continued living wherever they happened to be after the flood events subsided. With the availability of the many open niches and much organic material stirred up by the flood, foraminifers could have multiplied rapidly. Roth (1985) discussed the volume of biogenic sediments, especially foraminiferal oozes, on the ocean floor and showed that it is plausible that the foraminiferal deposits found on today's ocean floors could have been formed in the time since the flood.
CONCLUSIONS
The abundance, diversity and distribution of foraminifers in the fossil record exemplify many of the problems of fitting the fossil record into a short chronology, such as the multitude of species, large numbers of organisms, and apparent evolutionary sequences of simple to complex forms. Living species of foraminifers exhibit diverse morphological forms under varying environmental conditions, raising the possibility that many of the nominal species in the fossil record are actually ecophenotypes. In this case, a long time for evolution to take place would not be required. Presumed evolutionary sequences could then represent populations living in different environments or in the changing conditions during the flood and as it subsided.
Foraminifers are sparsely represented in the fossil record up through the Triassic, with most specimens found in small fossiliferous deposits. [?? 12 October 2001] With this in mind, and the fact that foraminifers can multiply rapidly, it seems plausible that the number of organisms found in the geologic column could have been produced in the time since creation.
The distribution of foraminifers in the fossil record seems to bear some resemblance to their ecological distribution in the oceans today. Fossil benthic species are found in most Phanerozoic strata, and similar forms are found living in ecological zones ranging from the deep sea to brackish estuaries. Fossil planktonic species are found only in Jurassic and younger strata, while living planktonic species are found floating in the upper parts of the water column. These distributions seem consistent with an ecological zonation model. The extinct large fusulinids in the Upper Paleozoic are anomalous, however. They are interpreted to have had symbiotic algae, so must have lived in shallow environments, perhaps in the upper ecologic zones of a low elevation sea.
REFERENCES
Bandy, O. L. 1953. Ecology and paleoecology of some California foraminifera. Part 1. The frequency distribution of Recent foraminifera off California. Journal of Paleontology 27:161-182.
Berger, W. H. 1976. Biogenous deep sea sediments: production, preservation and interpretation. In J. P. Riley and R. Chester (eds.). Chemical Oceanography, Vol. 5, 2nd ed., pp. 265-388. Academic Press, New York.
Bernhard, J. M. Characteristic assemblages and morphologies of benthic foraminifera from anoxic organic-rich deposits: Jurassic through Holocene. Journal of Foraminiferal Research 16:207-215.
Brasier, M. D. 1980. Microfossils. George Allen & Unwin, London.
Clark, H. W. 1946. The new diluvialism. Science Publications, Angwin, California.
Douglas, R. G. 1979. Benthic foraminiferal ecology and paleoecology: a review of concepts and methods. In J. H. Lipps, W. H. Berger, M. A. Buzas, R. G. Douglas, and C. A. Ross. Foraminiferal Ecology and Paleoecology, pp. 21-53. Society of Economic Paleontologists and Mineralogists Short Course No. 6.
Haynes, J. R. 1981. Foraminifera. Halsted Press, New York.
Hendrix, W. E. 1958. Phenotypic variation in some Recent and Late Cenozoic planktonic foraminifera. In R. H. Hedley and C. G. Adams (eds.). Foraminifera, Vol. 2, pp. 111-170. Academic Press, London.
Loeblich, A. R. and H. Tappan. 1964. Protista 2. In J. H. Lipps, W. H. Berger, M. A. Buzas, R. G. Douglas, and C. A. Ross. Foraminiferal Ecology and Paleoecology, pp. 54-61. Society of Economic Paleontologists and Mineralogists Short Course No. 6.
Roth, A. A. 1985. Are millions of years required to produce biogenic sediments in the deep ocean? Origins 12:48-56.
Schroder, C. J., D. B. Scott, F. S. Medioli, B. B. Bernstein, and R. R. Hessler. 1988. Larger agglutinated foraminifera: comparison of assemblages from central North Pacific and western North Atlantic (Nares Abyssal Plain). Journal of Foraminiferal Research 18:25-41.
Sliter, W. V. 1972. Cretaceous foraminifers depth habitats and their origin. Nature 239:514-515.
Tosk, T. A. and K. A. Andersson. 1988. Late Early Triassic foraminifers from dysaerobic to possibly anoxic environments of the Thaynes Formation, southeastern Idaho. Journal of Foraminiferal Research 18:286-301.