STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS
by Leonard Brand and James Florence Department of Biology Loma Linda University
WHAT THIS ARTICLE IS ABOUT
Fossil vertebrate footprints are distributed through much of the geologic column. Reptile and amphibian footprints are most abundant and diverse in Permian through lower Jurassic rocks. At the end of the lower Jurassic there is a sharp drop in diversity, and essentially the only footprints found in upper Jurassic and in Cretaceous rocks represent a few types of large dinosaurs. Vertebrate footprints are rare in the lower Tertiary, but mammal and bird tracks are more common in upper Tertiary rocks. The distribution of reptile and amphibian fossil footprints in the geologic column differ sharply from the distribution of reptile and amphibian fossil bones. Why are reptile and amphibian tracks so rare in upper Mesozoic and Tertiary deposits, in which the corresponding bones are abundant? It is suggested that these data are explained most naturally by an earth history model which includes a worldwide flood.
Fossilized footprints are a record of animal activities in the remote past. They can provide some types of information that fossil bones do not provide.
Fossil skeletons contain a wealth of information about animals that have become extinct long ago. If most of he skeleton is available, the general structure and appearance of the animal can be reconstructed with reasonable accuracy. From the characteristics of the teeth we can deduce something about the food habits of the animal. Bones and teeth can even provide much information beyond that, but can we be sure that the animal lived and died at the place where we found its fossil bones? In many cases the answer is no. When an animal dies its bones may be scattered by scavengers. If it dies near water it may be washed far downstream, or it may float in a body of water for some time before it sinks and is buried (Behrensmeyer and Hill 1980; Schafer 1972). After it is buried it may still not be able to rest in peace. At some later time the sediments in which it is buried may be eroded out, and the bones and sediments may be washed down to another basin and deposited again.
An animal's footprints in the mud cannot go through any of these processes and still be preserved. Consequently when we find a fossil animal footprint we know that an animal walked on that very spot at some time in the distant past. This is one of the characteristics of fossil footprints that makes them helpful in analyzing the history of life on earth. The study of fossil footprints and other fossilized evidence of animal behavior is called ichnology, and each type of footprint, believed to represent the tracks of one species of animal, is given a genus and species name and is called an ichnospecies.
METHODS
We quantitatively analyzed the stratigraphic distribution of fossil vertebrate footprints as revealed by data from about 800 published papers and from additional specimens in the American Museum, U.S. National Museum, Yale University Peabody Museum, and the Raymond Alf Museum. Much of the published footprint literature dates from the 19th and early 20th century, and contains considerable duplication several different workers giving different names to the same type of fossil tracks. As far as possible we avoided this excess of ichnospecies for reptiles and amphibians by following the taxonomy of Haubold's (1971) taxonomic revision, supplemented by more recent papers.
There is a large degree of uncertainty concerning the number of species of birds and mammals represented by fossil footprints, because of the difficulty of accurate identification and the lack of any comprehensive taxonomic study. Consequently in our graphs of bird and mammal footprint diversity we indicate maximum and minimum diversity that can be derived from the literature plus observed museum specimens.
In addition to the factors noted above, there are likely to be many undescribed specimens scattered in various museums that we have not seen. However, the sample reported here seems to be adequate to demonstrate reliable trends in distribution and diversity. The patterns illustrated in this paper were evident early in the data collection process, and further literature and museum study increased the numbers of taxa, but did not significantly change the relative trends.
We compared the diversity of fossil footprints with diversity of body fossils (bones and teeth) at successive stratigraphic levels. The body fossil data are from Harland (1967).
COMPARATIVE DIVERSITY OF FOOTPRINTS AND BODY FOSSILS
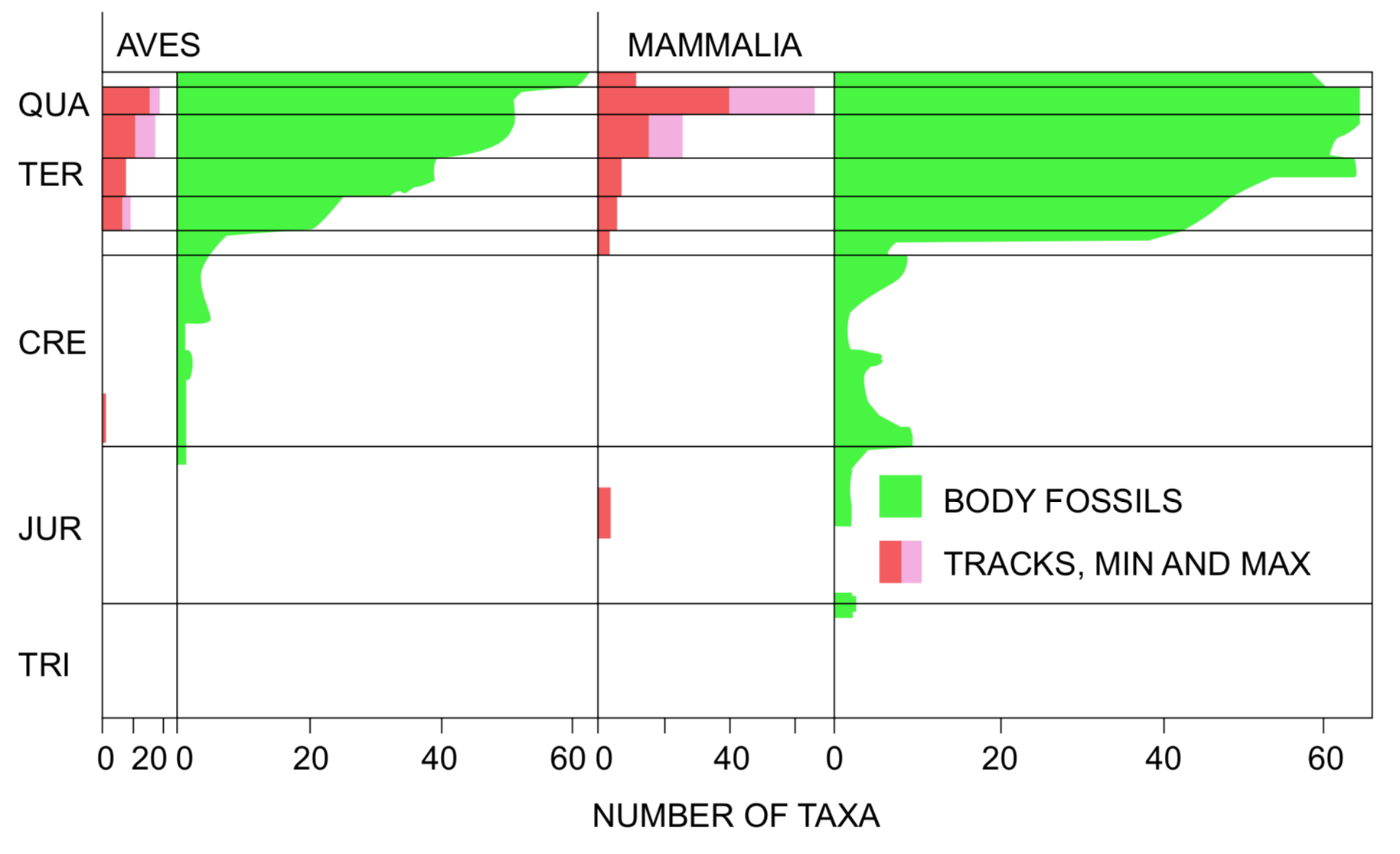
Overall distribution of bird and mammal footprints correlates well with the distribution of bird and mammal body fossils (Figure 1). Mammal footprints and bones are both most abundant in upper Tertiary deposits. The mammal footprint fauna includes one small mammal taxon in the Jurassic and one in the Miocene or Pliocene. The other mammal tracks are predominantly carnivores, ungulates (hoofed animals), and elephants. Bird footprints are never very diverse, and they are nearly limited to Tertiary deposits. The bird footprint fauna is dominated by shore bird types and large ground-dwelling birds. However, many are just identified in the literature as "bird tracks."
FIGURE 1. Comparison of diversity of fossil footprints and body fossils of birds and mammals. Footprint diversity is given in number of ichnospecies. Various taxonomic levels are represented in the body fossil data (from Harland 1967), with families being most common. Maximum and minimum figures for the number of footprint taxa indicate the range of uncertainty resulting from the incomplete taxonomic information for many of these fossils.
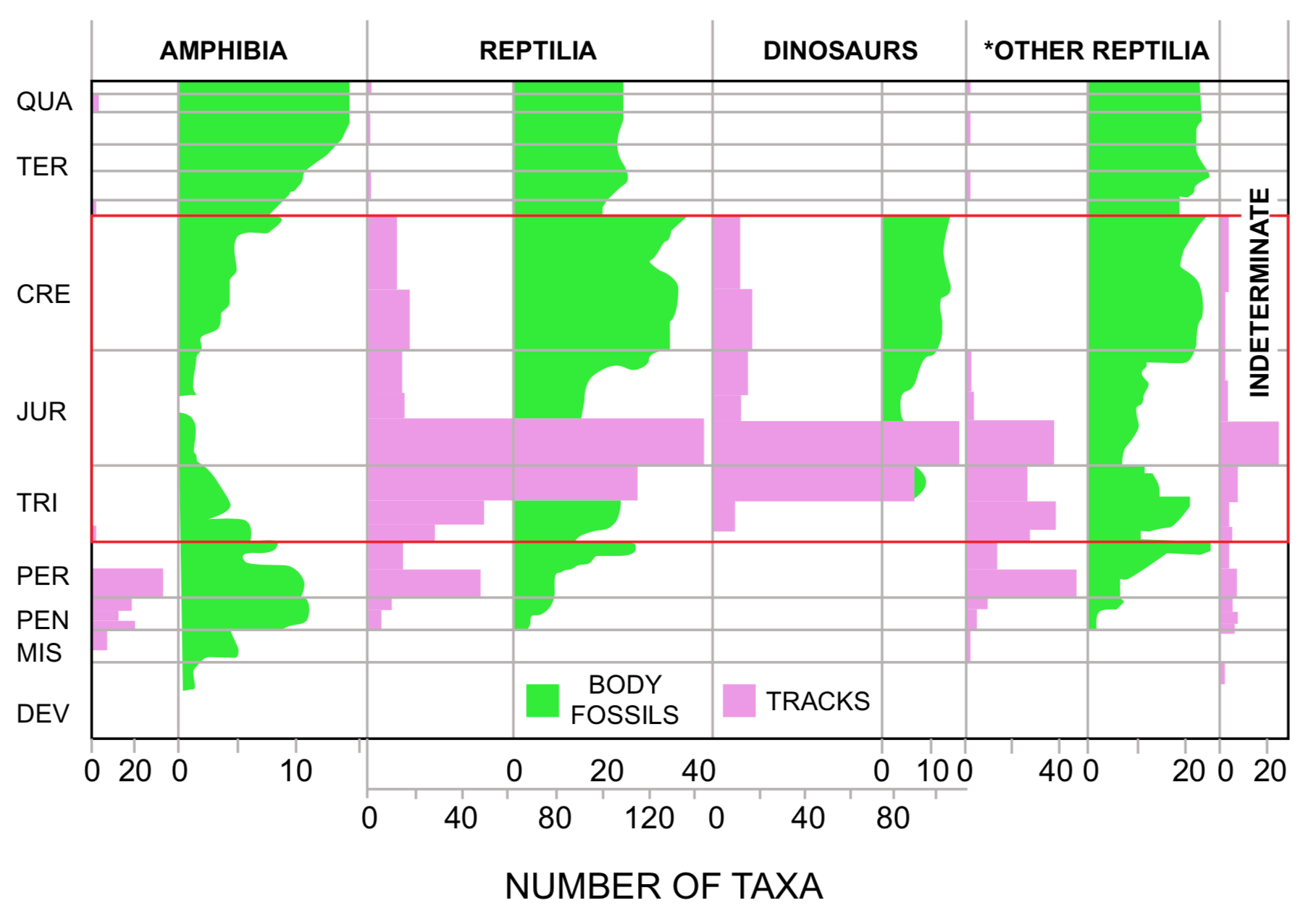
In contrast to the bird and mammal footprints, the stratigraphic distribution pattern of amphibian and reptile footprints is very different from the distribution of body fossils (Figure 2). Amphibian footprints are rare after the early Permian, and reptile footprints, with the exception of dinosaur footprints, which are the most diverse and conspicuous group of tracks, are most abundant in late Triassic and early Jurassic rocks (Figure 2). The only Cretaceous reptile footprints identified in the literature are about a dozen types of large dinosaur footprints. In contrast, amphibian and reptile body fossil diversity is greatest in the Cretaceous and Tertiary, when corresponding footprints are rare or nonexistent.
FIGURE 2. Comparison of diversity of fossil footprints and body fossils of amphibians, all reptiles, dinosaurs, and (*) reptiles exclusive of dinosaurs, and of indeterminate tetrapod footprint taxa. Footprint diversity is given in number of ichnospecies. Various taxonomic levels are represented in the body fossil data (from Harland 1967), with families being most common.
CONCLUSIONS
If the geologic column represents sediments that have accumulated over many millions of years, and the fossils from each geologic period are the remains of animals living in successive time periods, it would be reasonable to expect that the stratigraphic patterns of footprint diversity should roughly parallel the patterns of equivalent body fossil diversity the periods with the most kinds of dinosaur bones should have the most kinds of dinosaur tracks, for example. The bird and mammal fossil record fits that expectation quite well, but the reptile and amphibian record definitely does not. We will discuss two approaches to explaining this discrepancy.
The first approach assumes that much of the geologic column was deposited during a global flood. This model suggests that during the early to middle part of the flood large numbers of amphibians and reptiles were moving about, and thus producing footprints. Later during the flood (upper Jurassic and Cretaceous) there were very few live amphibians or reptiles to produce footprints, except for the large dinosaurs. During the Cretaceous when the only footprints preserved were the large dinosaur tracks, there were many amphibian and reptile bodies that were being buried to produce the abundant Cretaceous body fossils. During the Cenozoic almost no amphibian or reptile footprints were preserved.
This flood model suggests that during the flood the birds and mammals were in the uplands, away from the depositional basins, because of ecological differences and/or their more adaptable behavioral response to the unusual biological crisis caused by the flood. Consequently they left almost no footprints. This model further suggests that the upper Tertiary footprints were formed after the flood when geological processes were more like those observed today.
If this flood model is correct, and the birds and mammals were living contemporaneous with the Paleozoic reptiles, why aren't there at least a few bird or mammal tracks in Paleozoic sediments? It appears that there may be a few. In carboniferous deposits in Nova Scotia tracks were found that "superficially ... resemble the tracks of some of the wading birds, but of course there is little probability of their having been made by birds" (Sternberg 1933) (Figure 3). If these tracks had been found in Cenozoic deposits it seems likely that they would have been described as bird tracks. Another interesting track was found in the Permian Hermit Shale of the Grand Canyon (Gilmore 1927). It looks precisely like a bird track, but since birds are not thought to have evolved until the Mesozoic, this Permian track is just listed as an "unidentified track" (Figure 4).
FIGURE 3. Unidentified, bird-like fossil footprint from Paleozoic strata. A trackway from the Carboniferous of Nova Scotia (after Sternberg 1933).FIGURE 4. Unidentified, bird-like fossil footprints from Paleozoic strata. A track from the Permian Hermit Shale of the Grand Canyon (after Gilmore 1927).
Another question that arises is why reptile and amphibian footprints are so abundant in Paleozoic and lower Mesozoic sediments and so rare in recent sediments, since reptiles and amphibians are common today. Part of the answer could be that there were many types of reptiles and amphibians living earlier in earth history that no longer exist. We would suggest that another part of the answer is that the rapid sedimentary processes during the global flood were usually conducive to the burial and preservation of footprints before they were destroyed by weathering processes. Thus during the flood there would have been a great diversity of reptiles and amphibians making tracks under conditions uniquely suitable for preserving those tracks. In more recent times, with most of the Mesozoic track-makers extinct and conditions not as suitable for preserving footprints, reptile and amphibian footprints seem to be seldom preserved.
The second approach to explaining these data is based on the conventional geologic model of sedimentation over long ages of time, coincident with the evolution of animal life. This model must assume that the stratigraphic distribution of amphibian and reptile fossil footprints is an artifact, and does not at all reflect the actual relative amount of animal activity during different geologic periods. Several factors have been suggested as potential contributors to producing this artifact:
1) In Mesozoic and Tertiary deposits containing larger, more conspicuous tracks, smaller tracks may be more likely to be overlooked, and not collected. This argument is weakened by the fact that small reptile tracks are abundant in Triassic and Early Jurassic rocks, when dinosaur tracks are also at their peak of abundance. It also does not explain the near absence of amphibian and reptile tracks in Early Tertiary deposits, which have no dinosaur tracks and few bird and mammal tracks to divert attention from the smaller tracks.
2) Smaller, shallower tracks would be destroyed more easily by weathering or by slumping of waterlogged sediment than the larger tracks. However, this factor should have affected the entire geologic record, not just the post-Jurassic deposits.
3) Older rocks tend to be well-indurated (firmly cemented), while younger rocks are more likely to be relatively unconsolidated, and thus less suited to preserve footprints. However, many of the Cenozoic footprints are quite well-preserved, including delicate bird tracks. Also this factor does not explain the sharp drop in footprint diversity after the lower Jurassic.
These factors may play a role in biasing the published footprint record, but they do not seem adequate to explain the sharp contrast between the abundant footprints of amphibians and small reptiles in Permian to Lower Jurassic rocks, and their near absence in younger rocks.
The only explanation that this model provides for the bird-like Paleozoic tracks is that they must have been made by unknown types of Paleozoic reptiles with bird-like feet.
We conclude that these data are most easily explained by the global flood model. Does this mean that they should be taken as proof of the flood? No, it does not mean that. Science rarely makes its decisions on something that could be called proof, but it makes its decisions on the perceived weight of evidence. This is particularly true when studying unobservable events from the past history of our earth. Our footprints can neither prove nor disprove the flood model. The question is, into which model do the data make the most natural fit? It appears that the footprint data make a very natural fit with a global flood model.
REFERENCES
Behrensmeyer, A.K. and A.P. Hill. 1980. Fossils in the making. University of Chicago Press. 338 pp.
Gilmore, C.W. 1927. Fossil footprints from the Grand Canyon. Second Contribution, Smithsonian Miscellaneous Collections 80(3):1-78.
Harland, W.B., ed. 1967. The fossil record. Geological Society of London. 827 pp.
Haubold, H. 1971. Ichnia amphibiorum et reptiliorum fossilum. In O. Kuhn, ed., Handbuch der Palaoherpetologie, Pt. 18, 124 pp. Gustav Fischer Verlag, Stuttgart and Portland, Oregon.
Schafer, W. 1972. Ecology and palaeoecology of marine environments. University of Chicago Press. 568 pp.
Sternberg, C.M. 1933. Carboniferous tracks from Nova Scotia. Geological Society of America Bulletin 44:951-964.
Front Cover: Numerous trackways from the Permian De Chelly Sandstone near Monument Valley, Arizona. See Brand-Florence article on p. 67. Back Cover: Close-up of amphibian or reptile trackway from the De Chelly Sandstone.