THE ORGANIC LEVELS OF THE YELLOWSTONE PETRIFIED FORESTS
by Harold G. Coffin Geoscience Research Institute Berrien Springs, Michigan
WHAT THIS ARTICLE IS ABOUT
In the fossil forests of Yellowstone one sees scores of layers of petrified trees (many of them upright) that have been interpreted as successive forests preserved in growth position. Many thousands of years would be required to grow these successive forests; hence, they have significant time implications for the creation and evolutionary models of origins. At the base of many of the tree layers, one finds finer sediments that contain organic material and that have been interpreted as soils. One may assume either the trees grew in their present position or that trees and soil were transported and deposited about the same time. A sequence of successive forests could be accommodated within a short period of time by the transport model which is favored by the author who presents evidence indicating that the organic zones do not show normal soil features.
The main points in support of this view are: 1) The organic zones are often thin or absent despite the presence of abundant or large petrified trees. 2) The organic profile may be reversed to that expected in normal soils, and there is no evidence of differential decay from top to bottom. 3) There is no evidence of the presence of animals or animal activity as would be expected on a normal forest floor. 4) Several areas show complex multiple and branching organic levels which suggest transport rather than in situ development. 5) Vertical microscopic sections through the organic layers show the type of sedimentary features and sorting of organic material expected from transport. 6) Clay is usually absent or, when present, does not appear to be related to the organic zones. 7) Un- weathered feldspar crystals which are expected from rapid burial and not from slow soil development are abundant.
Introduction
The Yellowstone petrified forests of Wyoming and Montana consist of scores of superimposed sequences of petrified trees. Because many of these trees are preserved in an upright position, the sequence gives the appearance of one forest having grown above another. Many thousands of years would be involved in the growth of these successive forests; hence, this topic is of considerable interest to various time-related interpretations of earth history.
Much of the recent discussion (Ritland and Ritland 1974, Coffin 1979) of these forests centers on the question of their origin autochthonous or allochthonous in situ growth or transport. The latter proposal would tend to negate the thousands of years that would be required for the growth of successive forests. One aspect of this question relates to the nature of the fine sediment layers containing organic material often found at the base of the trees. Do these layers show time-dependent maturing characteristics as expected in true soil development, or do they show evidences of rapid transport, thus not implying much time for formation?
Much of the research on the Yellowstone petrified forests since the first report by Holmes (1878) has been taxonomic. Knowlton's monograph (1899) was the first and most extensive. Read (1933) and Dorf (1960) have examined and revised the work of Knowlton. Dorf also discussed the paleoecology and interpreted the organic zones as growth surfaces or pockets and lenses of organic debris deposited by small streams. Ritland and Ritland (1974) and Coffin (1979) have discussed basic models of formation, and Fritz (1979) discusses various depositional features. Beyond this, little attention has been given to the organic zones except as sites for the collecting of specimens for taxonomic study.
The fossil forest areas most frequented lie along the slopes and cliffs of Specimen Ridge which flank the Lamar Valley on the south. Two classic areas generally referred to as Specimen Ridge and the Fossil Forest lie several miles apart opposite Slough Creek and Soda Butte Creek respectively. However, fossil forests also exist in and beyond the northwest corner of the Park, on both sides of Soda Butte Creek, and on the east side of Cache Creek. Other less significant sites are known in the northern and eastern parts of the Park. In addition, an extensive area exists in the Stratified Primitive Area south of Yellowstone Park. The research reported in this paper has been undertaken in all the sites mentioned above except the Stratified Primitive Area. All these petrified forests are in Eocene volcanic breccia and ash beds.
General Description
The upright trees (Figure 1), which sometimes reach as high as 6.7 m and occasionally even higher, have a diameter up to 4.5 by 3.8 m and are often seen to arise from layers or zones which are composed of needles, leaves and organic debris. These levels lie in positions in relation to the roots of the upright petrified trees that correspond to growing surfaces upon which humus and soil have accumulated (Figure 2). They have been interpreted as soil levels a natural and obvious interpretation.
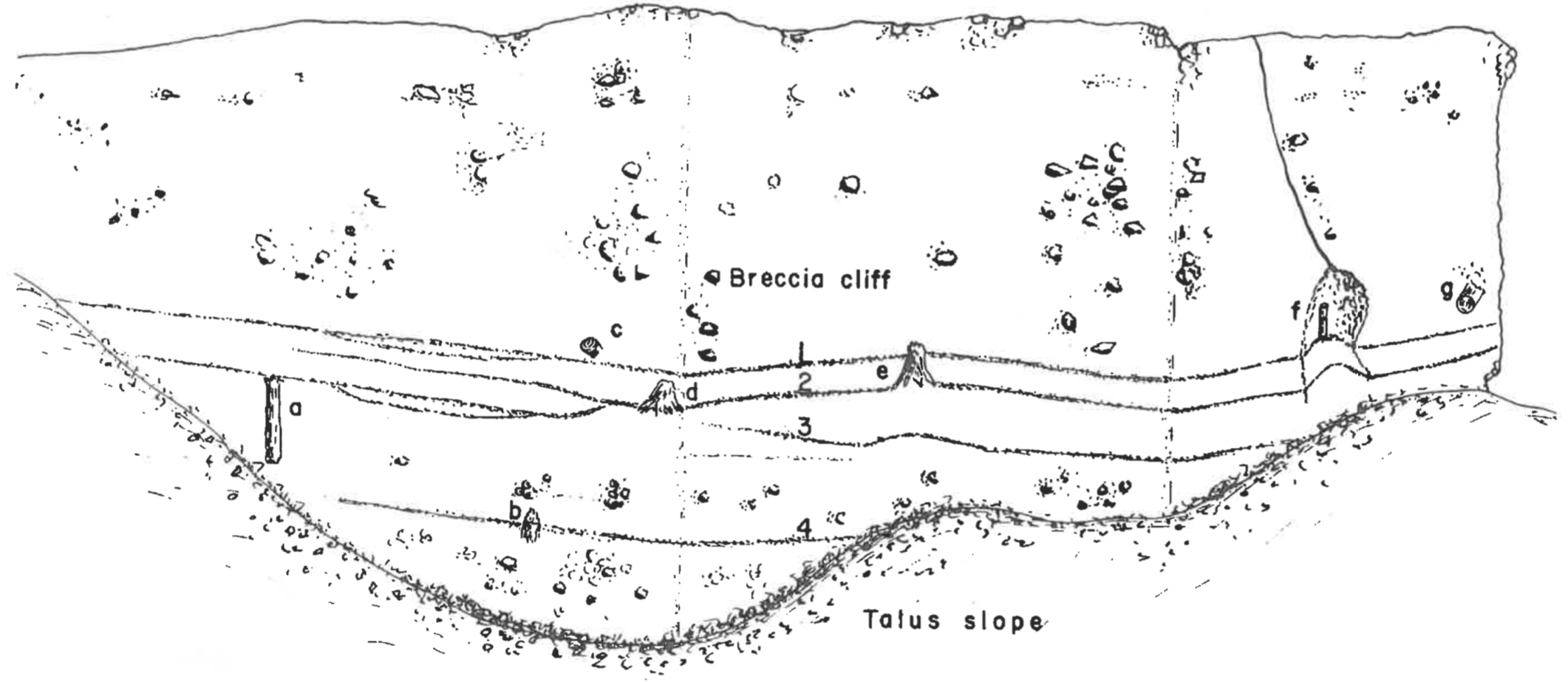
FIGURE 1. Vertical broomstick-size petrified tree in volcanic breccia, Mt. Norris, Yellowstone National Park. The tree is to the right of the picture and is about 1.5 m high.FIGURE 2. Sketch of a breccia cliff in the Cache Creek petrified forest showing a complex arrangement of organic levels and trees.
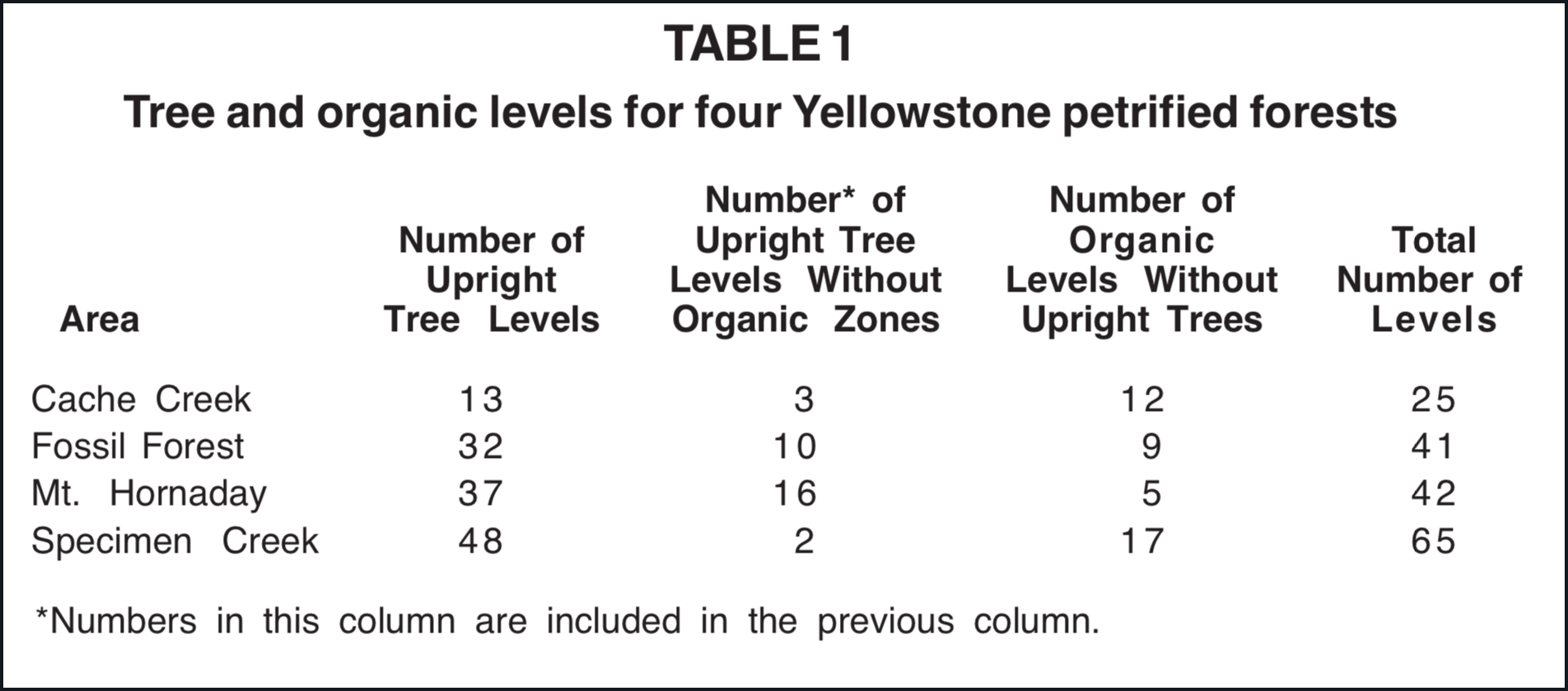
However, many of the organic levels of the Yellowstone petrified forests are thin and contain only a small fraction of the organic material that would be expected, based on the sizes of the trees arising from the levels. Sixty-three levels on the slopes above Specimen Creek range from a trace of organic matter to 15 cm thick. The average is close to 3 cm. These dimensions are for the total depth of "soil" or organic matter. A distinction between forest floor litter and underlying soil is not visible. Of the 130 levels with upright trees included for the four petrified forests listed in Table 1, 24% contain no organic matter; however, the situation appears highly variable. Sixteen of the 37 levels of Mt. Hornaday have no forest debris whereas only two of the 48 levels of Specimen Creek are exceptions.
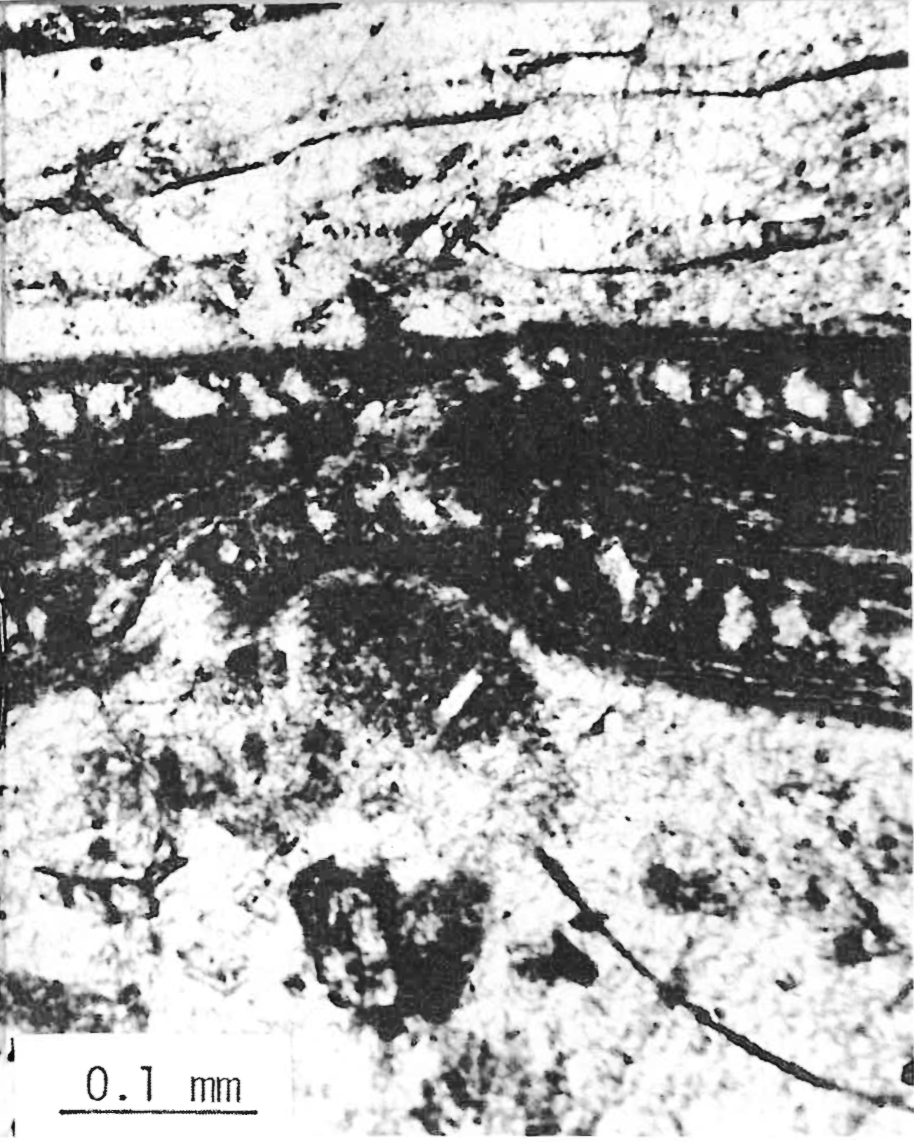
Preservation of the organic matter is excellent. This good condition has, of course, facilitated the identification of the plants. Occasionally even the detailed cellular structure of specific tissues of leaves and needles can be seen in thin-section slides (Figures 3 and 4).
FIGURE 3. Photomicrograph of petrified deciduous leaf in volcanic ash from Mt. Hornaday, Yellowstone National Park.FIGURE 4. Photomicrograph of the cross-section of a petrified coniferous needle from Cache Creek petrified forest.
Evidences of Reworking by Water
Cross-sections through true soils reveal a typical profile of organic density resulting from increased blackness or richness of humus toward the surface of the ground. Eighty-six (71.6%) of 120 vertical thin sections of organic levels studied have the organic matter mixed into the sediments with no prevailing order of density. Twelve of the sections (10%) have a reverse organic profile with the greatest accumulation of organic matter at the bottom.
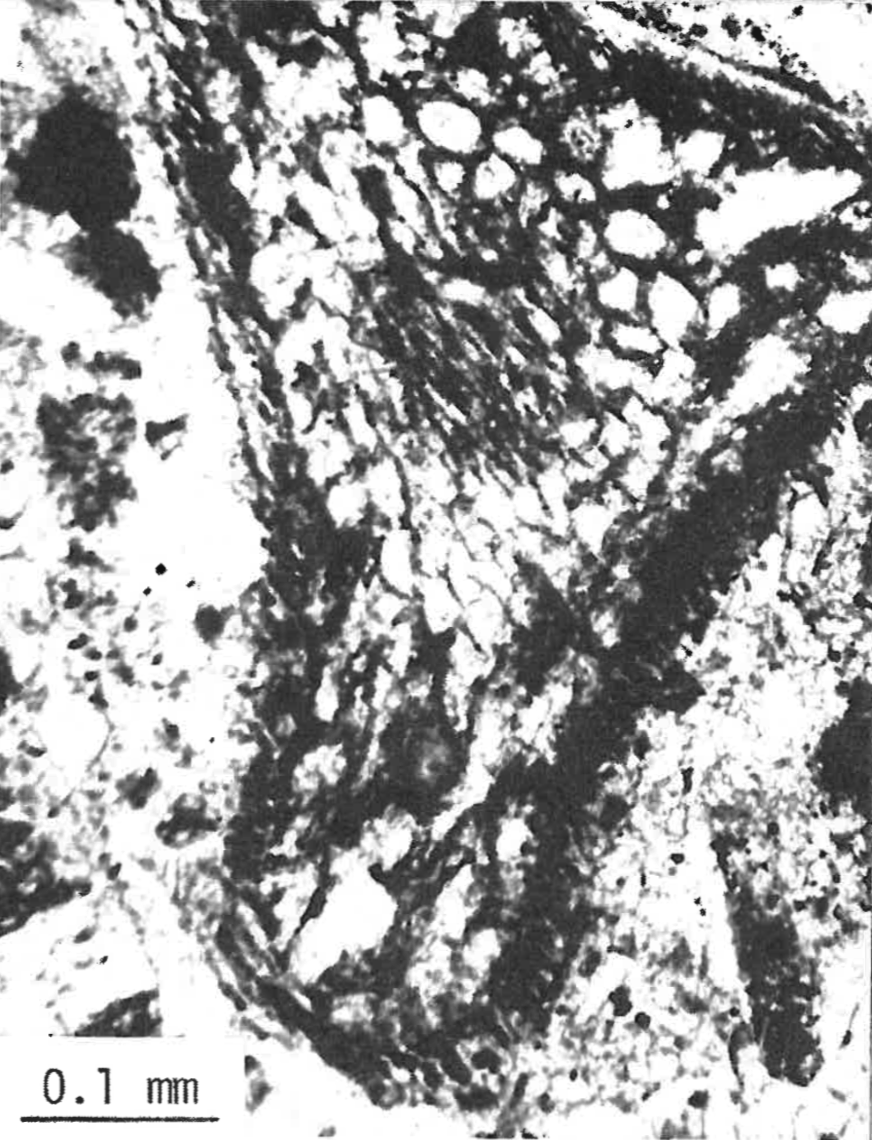
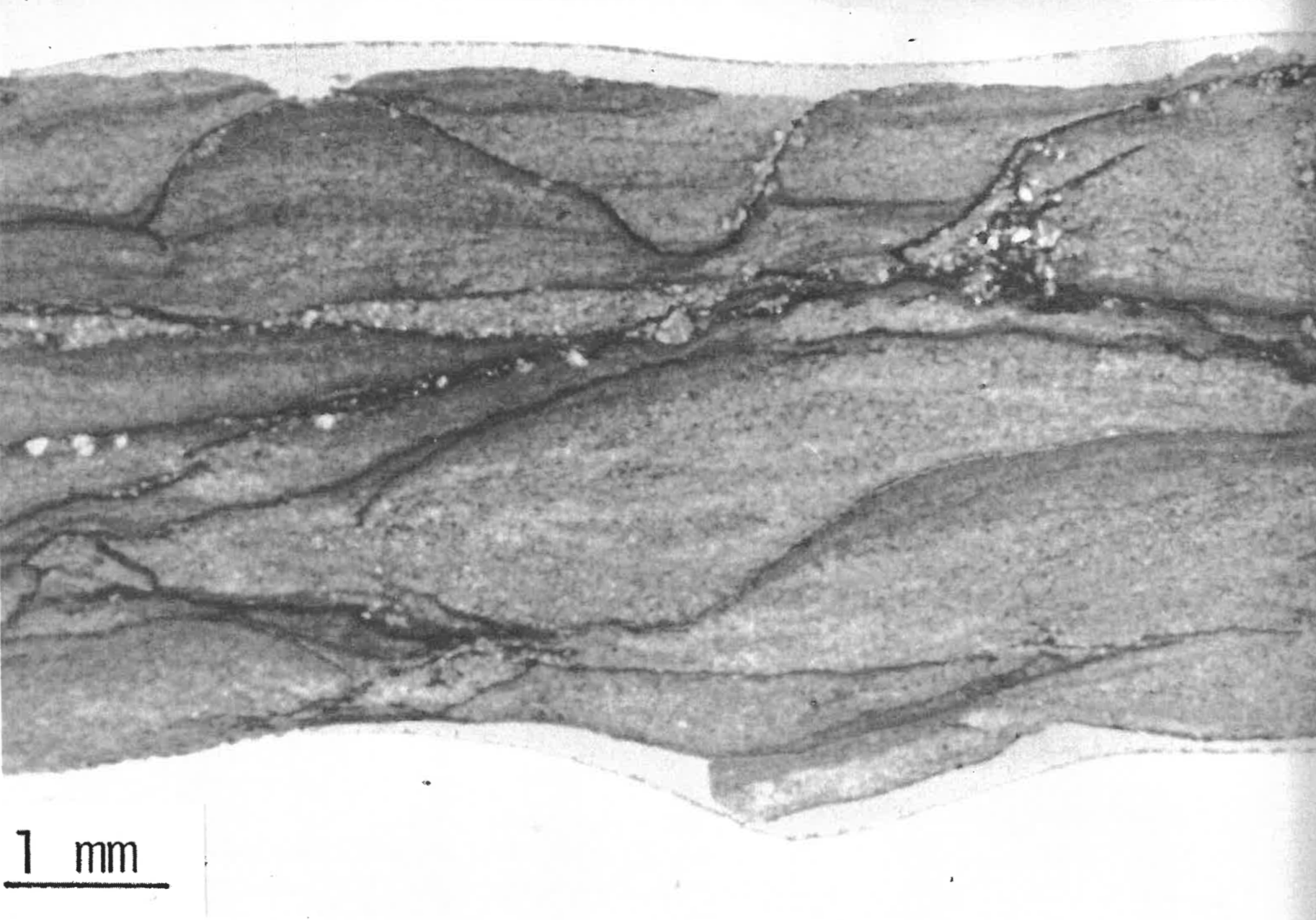
There is also sorting of organic material in 20% of the sections from 19 levels in the Specimen Creek Fossil Forest, Mt. Norris and Miller Creek petrified forests. Figure 5 shows a relationship between the size of the ash sediment and the size of the organic material fine sediment, fine organic matter; coarse sediment, coarse organic matter. Similar to this example is size sorting of the inorganic particles among or between leaves. Figure 7 is from an Oregon site, but also illustrates the Yellowstone situation. The leaves are seen in cross-section as long, somewhat undulating lines. Between the lines the sediments show normal grading (also see Coffin 1979, Figure 8).
FIGURE 5. A vertical thin section of an organic level from Mt. Norris. Note the sorting of both organic and inorganic matter. The dark streaks and spots are vegetable matter.FIGURE 7. Enlargement of a vertical thin section from an organic level near Cascade Locks, Oregon. The petrified tree-volcanic breccia relationships here are similar to those of Yellowstone. Note the normal grading of sediments between the deciduous leaves, seen here in cross-section.
In a normal soil, undecayed leaves and needles are confined mostly to the surface. Material more than a few seasons old disintegrates into humus, and identification becomes difficult or impossible. Differential decay with depth is absent in the organic levels of Yellowstone. Throughout the petrified forests of the region, vegetable debris at the bottom of a layer is as well preserved as that at the top.
A feature known for many years is the absence of animal remains. Volcanic activity could have caused larger forest animals to flee elsewhere. This explanation is not satisfactory, however, because many animals could not or would not leave their forest habitats. Land snails, some amphibians and reptiles, many insects, arachnids and worms would not escape burial. Eggs and young of many types would be unable to flee. In addition, bones, teeth, scales, exuviae, castings, droppings, burrows, etc., would qualify as evidence of animal life. None of these have been found in the organic levels during a century of research. Considering the excellent preservation of the delicate plant parts, diagenetic destruction of animal remains appears unlikely.
Complexity of Organic Levels
The complexity of the organic levels is apparent especially in the Cache Creek and Specimen Creek petrified forests. Figure 2 is a sketch of a section of cliff in the Cache Creek area. Notice how some of the organic levels split and recombine. Levels one and two are less than a meter apart. Note the penetration of trees b and e through overlying organic zones. Even more complex are the Specimen Creek organic levels (see Coffin 1979, Figure 6).
Could the upper organic bands of multiple levels represent the leaf-fall zones associated with air-drop ash in volcanic eruptions? In such cases the lowest band would represent the true soil level, whereas the upper one(s) would result from physical and chemical stripping of leaves and needles from the trees by explosive volcanic activity. These upper bands should not be growth surfaces unless no further ash accumulation occurred for many years and a new forest established itself on this level. The study accompanying the survey of the complex Specimen Creek levels failed to distinguish any significant differences between surfaces from which visible trees arise and adjacent bands containing no visible upright trees. Growth levels with trees and also bands within levels were sampled and examine in the thin section studies described earlier. If leaf-drop zones are present, they are not readily apparent and cannot be distinguished from the other levels. Until some quantitative feature for separating leaf-drop from growth levels is found, evaluation of this possibility is difficult.
Taxonomic Characteristics
In this research gross identification of the plant specimens in the organic levels was undertaken. Trees were classified as Pine-type (resin ducts present), Sequoia-type (no resin ducts), and deciduous (vessels present). Leaves and needles in the organic levels were identified to the same categories.
Taxonomic sorting of the constituents in the organic bands were noticed early in the research. It resulted in a sequence of broad leaves at the top of the organic zone, mixed broad leaves and needles just below, and only needles at the bottom. Under normal conditions leaves, needles, cones, limbs, bark, etc., fall as a well-mixed litter onto the forest floor year by year as the seasons pass and the trees grow. A flotation experiment involving aspen and poplar leaves and fir needles in a tank of water showed that the needles became saturated and sank to the bottom first. Thus flotation in water is a possible explanation for this kind of taxonomic sorting.
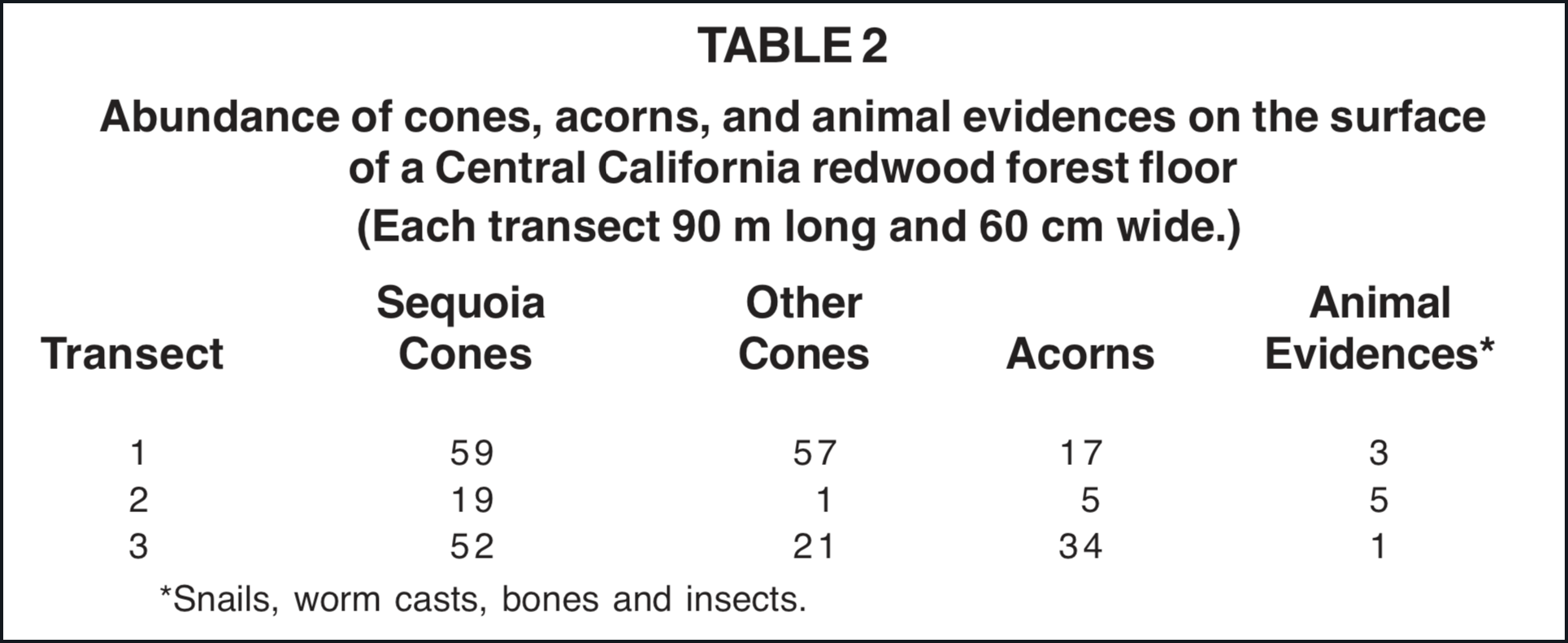
Three transects 90 m long and 2/3 m wide in a central California mature redwood forest showed Sequoia and other cones on the surface of the forest floor (Table 2). Although Sequoia cones are small and fragile, they do remain intact and visible for several months after failing. No petrified Sequoia cones have been found in Yellowstone despite the dominance of Sequoia trees. Cones of any type are rare in the petrified forests.
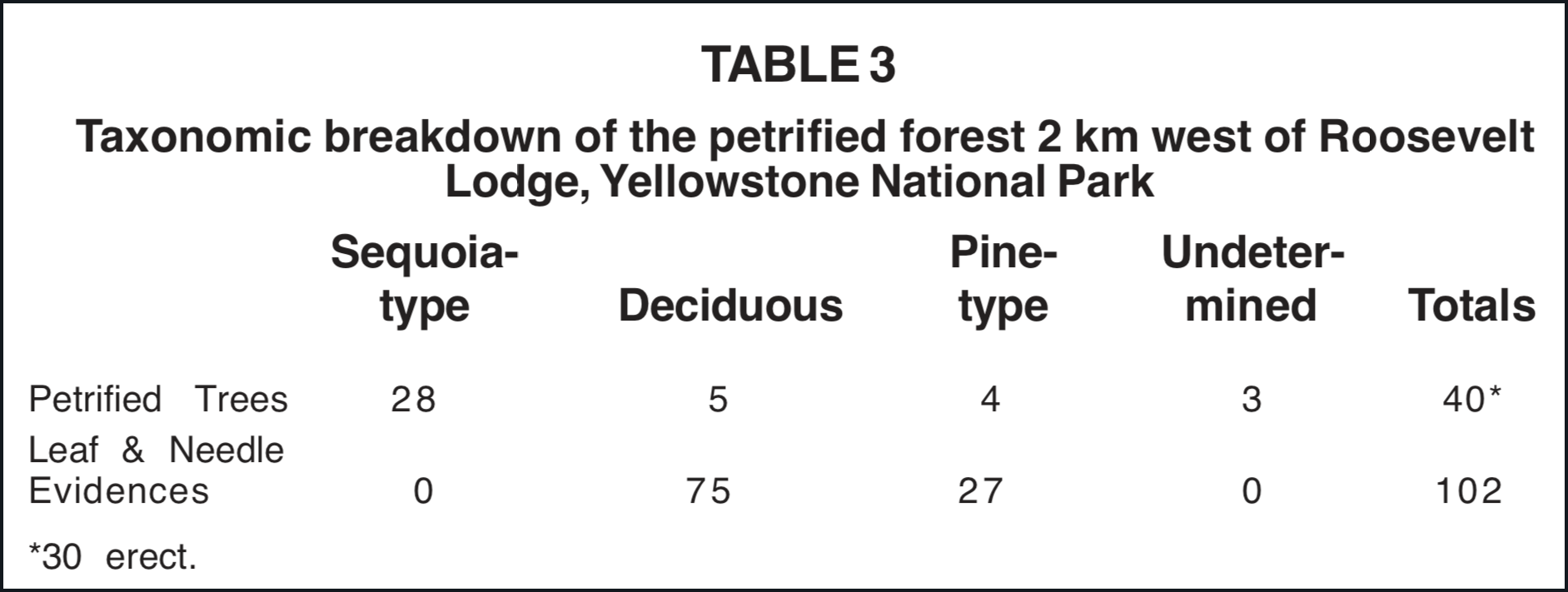
In a mixed forest of redwood and deciduous trees such as exists in California, the redwood needles greatly predominate in the forest floor litter. For the area overshadowed by a tree, conifers appear to drop proportionately many more needles than do deciduous trees their broad leaves. Knowlton (1899, p. 757) remarked about the absence of needles in the organic levels associated with the fenced petrified tree near Roosevelt Lodge in Yellowstone National Park. Our studies there have been summarized in Table 3. There is a lack of taxonomic agreement between the dominant trees in the area and the leaf and needles in the organic layers. One would expect to find great numbers of Sequoia needles and some cones, since most of the upright trees are Sequoia. However, large numbers of broad leaves and only a few pine needles are seen in the organic levels. Sequoia needles were entirely absent.
Many genera are represented only by pollen, but this might be expected, since the wood samples have not received thorough taxonomic study. More difficult to explain, if the trees are in position of growth, are the cases represented by wood or leaves only. Trees with wind-transported pollen such as walnut and sycamore should have left a pollen record in the forest floor, but little or no pollen for these two has been found.
DeBord (1977) worked in the petrified forest in the northwest corner of the Park (Specimen Creek). Four levels especially were given careful analysis. Pollen obtained from samples taken from 100 meter sections for each of the four levels were compared within levels, between levels, and between micro-and megaflora. Modern forest floors contain pollen in abundance inversely proportional to the distance from the source trees especially for trees using wind as the pollen-transporting force (Tauber 1965 and 1976, Anderson 1970 and 1973). No positive correlation exists between Yellowstone fossil pollen abundance and the proximity of possible source trees. The differences between individual samples on the same level are such that single sample analysis cannot be used to adequately describe the level.
The taxonomic composition of any particular forest of trees should influence the composition of the next higher forest of the area (if the trees are in growth position) because in most cases only the lower trunks of the standing trees would be covered by the advancing breccia-mud slide. The cones, seeds, nuts, and fruits would fall from the upper parts of the partially buried trees and repopulate the new surface with a similar forest. In DeBord's study (1977) no positive correlation exists between the taxonomic composition of pollen of the organic levels of one forest with that of forests directly above or below. Pine pollen was under-represented in three of the four forests analyzed. One of these three levels showed a severe under-representation of pine pollen and a severe over-representation of deciduous pollen.
Other Related Evidence
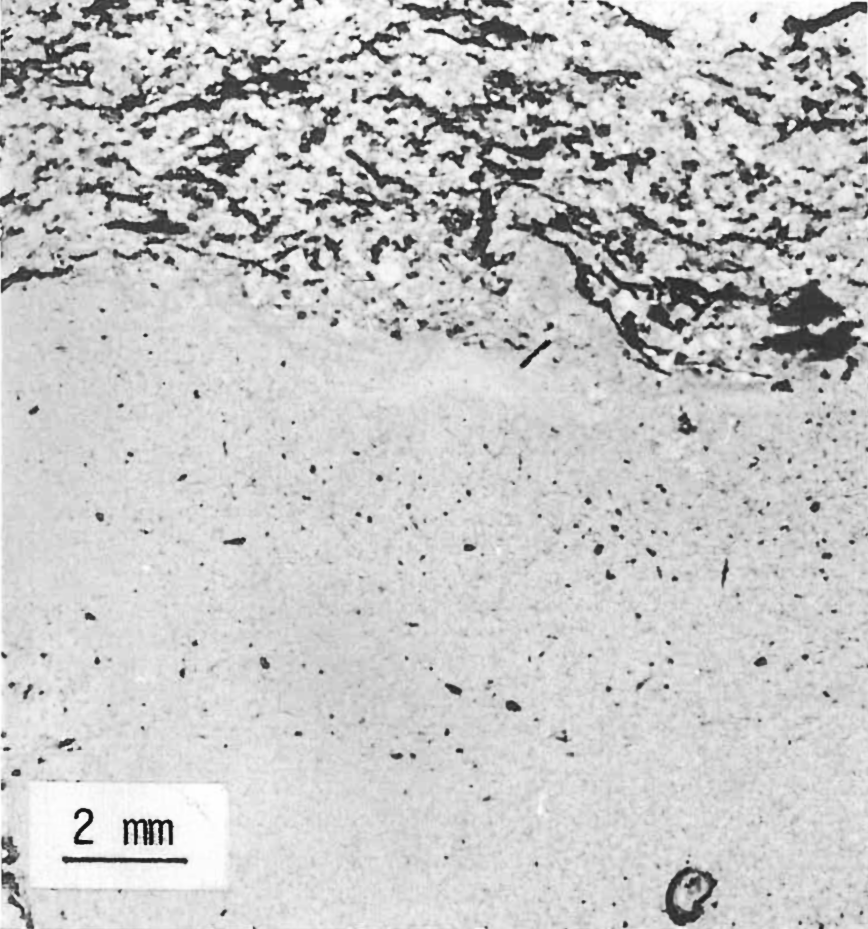
The presence of a clay profile formed by the slow breakdown of minerals is related to normal soil maturity processes. An analysis by x-ray diffraction and infrared scans on over 350 samples has been done for the Specimen Creek Area. Of nine horizontal bands of clay (montmorillonite) found distributed through the Specimen Creek petrified forest, three were limited to the breccias between organic levels. Five included one organic level and portions of the breccia beds immediately above and below and one included two organic levels and associated breccia beds. Clay content was up to 60% but no profile was detected on any of the seven organic zones included in the clay bands. Horizontal sampling of two clay bands at 5 to 10 feet intervals for 100 feet showed a constant mineral makeup. Abundant unweathered feldspar is scattered throughout the Yellowstone organic levels (Figure 6), suggesting rapid burial.
FIGURE 6. Unweathered crystals of plagioclase from an organic level on Mt. Hornaday. Note arrows.
None of the other organic zones contained detectable amounts of clay. Clay detection limits were at the 1-2% level. The apparent absence of clay in the majority of levels raises temporal questions concerning the so-called soil zones. Furthermore the sudden abundant appearance of clay in a few horizontal bands that include both organic levels and breccia beds suggests transport rather than in situ formation.
The rate of clay formation is variable, depending on climate and the parent rock. A sequence of mud slides on Mt. Shasta ranging in ages from 27 to 1200+ years showed little increase in clay content with age (Dickson and Crocker 1953). In contrast, clay formed on the volcanic ash soils of the West Indian island of St. Vincent at the rate of 1½ to 2 ft./1000 years (Hay 1960). Some levels in Yellowstone with large trees (up to 15 feet in diameter) would represent soil development well over 1000 years duration if the trees are in growth position. The mixed flora of the Yellowstone fossil forests makes it difficult to determine what the past climate of the region would have been.
The tentative results from the study of clay appear to suggest that no significant passage of time has been involved in the formation of the organic levels of Yellowstone. Spark source mass spectrometry research (in progress) appears to give the same results. The lack of significant difference between organic levels points toward these levels (and associated breccia beds) as being the result of one rapid volcanic episode.
Comparisons with Other Breccias
Fiske (1963, pp. 391-406) has described volcanic lahars of the extensive Ohanapecosh formation in Mt. Rainier National Park, Washington. In these breccias we found some organic levels, although they are less strongly developed than those of Yellowstone. Horizontal petrified trees also are noted. Fiske interprets these breccias as being subaqueous deposits. Obviously these organic levels cannot be growth surfaces if the deposits slid into position under water.
A road cut for Interstate 80 in the Miocene Eagle Creek formation near Cascade Locks, Oregon, exposed several levels of petrified trees and organic debris in volcanic breccia a situation closely similar to that of Yellowstone. Whatever interpretation is achieved for the Yellowstone breccias will probably apply to the breccias of this Oregon location and vice versa. Figure 7 is an unusual section of the organic level of the one remaining vertical tree still visible in the road cut. The pronounced normal gradation of sediments between the deciduous leaves (seen in cross-sections) is unique. Such grading hardly could be produced in normal undisturbed soil and suggests transport.
Conclusion and Implications
The normal accumulation of organic debris and the subsequent formation of humus and true soil which proceeds relentlessly on modern growth surfaces does not readily account for several of the phenomena seen in the organic levels of Yellowstone. Specifically these are:
The thinness or absence of organic matter on levels with abundant and large trees.
Good preservation no differential decay from top to bottom.
The absence of evidences of animals expected in typical forest plant-animal associations.
The multiplicity and complexity of the organic levels.
Lack of agreement between organic components and the dominant tree types as seen by studies of leaf types and pollen.
The evidences of contemporaneous water deposition of sediments and organic matter seen in the thin sections of the organic levels.
The vertical sorting of organic matter and atypical soil profiles.
The absence of weathering of feldspar crystals in many organic levels.
The absence of anomalous arrangement of clay in the organic levels.
The movements of volcanic lahars over the ground could eliminate a normal soil profile but they would not be expected to produce a reverse profile or sort the organic matter. These phenomena might be produced by small streams sorting and redepositing humus and forest litter as suggested by Dorf (1960, p. 257). However, these organic levels are often widespread and uniform in thickness. This feature and the absence of scouring or erosion would appear to eliminate small streams as agents for sorting and redepositing organic matter. Widespread flooding associated with volcanic activity and preceding each breccia-mud slide might be responsible for some of the anomalies seen in the organic levels. Such water activity could be responsible for features described in items 1, 2, 4, 6, 7, 8, and 9 above. The absence of animal remains and the lack of agreement between micro- and macroflora (items 3 and 5) suggest transport and sorting in a sea or large lake.
The unusual problems posed by the Yellowstone Petrified Forests and their surrounding sediments challenge research from multiple disciplines.
I wish to acknowledge the work in palynology (Lanny H. Fisk and Phillip L. DeBord), surveying and plotting (Donald G. Jones), and geochemistry (Ivan G. Holmes and Clyde Webster, Jr.) that I have cited in this paper.
REFERENCES
Anderson, S.T. 1970. The relative pollen productivity and pollen representation of North European trees, and the correction factors for the pollen spectra. Geological Survey of Denmark, Series No. 96.
Anderson, S.T. 1973. The differential pollen productivity of trees and its significance for the interpretation of a pollen diagram from a forested region. In H.J.B. Birks and R.E. West, eds. Quaternary Plant Ecology, pp. 108-114. Blackwell Scientific Publications, Oxford.
Coffin, H.G. 1979. The Yellowstone petrified forests. Spectrum 9(4):42-53.
DeBord, Phillip L. 1977. Gallatin Mountain "petrified forest": A palynological investigation of the in situ model. Ph.D. dissertation, Loma Linda University.
Dickson, B.A. and R.L. Crocker. 1953-1954. A chronosequence of soils and vegetation near Mt. Shasta, California. I, II, III. Journal of Soil Science 4:123-154; 5:173-191.
Dorf, Erling. 1960. Tertiary fossil forests of Yellowstone National Park, Wyoming. Billings Geological Society, 11th Annual Field Conference Guidebook, pp. 253-259.
Fiske, Richard S. 1963. Subaqueous pyroclastic flows in the Ohanapecosh formation, Washington. Geological Society of America Bulletin 74:391-406.
Fritz, William. 1979. Depositional environment of the Yellowstone fossil forests as related to Eocene plant diversity. Geological Society of America Abstracts 11(7):428.
Hay, Richard L. 1960. Rate of clay formation and mineral alteration in a 4000-year-old volcanic ash soil on St. Vincent, B.W.I. American Journal of Science 258:354-368.
Holmes, W.H. 1878. Fossil forests of the volcanic Tertiary formations of the Yellowstone National Park. U.S. Geological and Geographical Survey Territorial Bulletin 5:125-132.
Knowlton, F.H. 1899. The Tertiary floras of the Yellowstone National Park. U.S. Geological Survey Monograph 32:651-691, Plates 77-121.
Read, C.B. 1933. Fossil floras of Yellowstone National Park, Part 1, Coniferous woods of Lamar River flora. Carnegie Institution of Washington Publication 416:1-19.
Ritland, R.M. and S.L. Ritland. 1974. The fossil forests of the Yellowstone region. Spectrum 6(1-2):19-66.
Tauber, H. 1965. Differential pollen dispersion and the interpretation of pollen diagrams. Geological Survey of Denmark 11, Series No. 89.
Tauber, H. 1967. Investigations of the model of pollen transfer in forested areas. Review of Paleobotany and Palynology 3:277-286.
COVER PICTURE. View of part of the buried fossil forests in the Specimen Creek area of Yellowstone National Park. Several vertical fossilized trees appear as lighter stumps and logs among the layers of volcanic breccia and tuff. The article by Dr. Harold Coffin in this issue discusses some of the time implications of this geologic feature.