El 10 de septiembre de 2015 un artículo de acceso abierto en la revista eLife (descargable en http://elifesciences.org/content/4/e09560) presentó a la comunidad científica la última incorporación de una especie al registro fósil de homínidos: Homo naledi (Figura 1)[1]. Fue una puesta en escena bien orquestada del descubrimiento que incluyó el lanzamiento de un documental de casi dos horas de duración producido por PBS y National Geographic titulado "Dawn of Humanity" y la clásica amplificación por parte de los principales medios de comunicación del sensacional hallazgo.
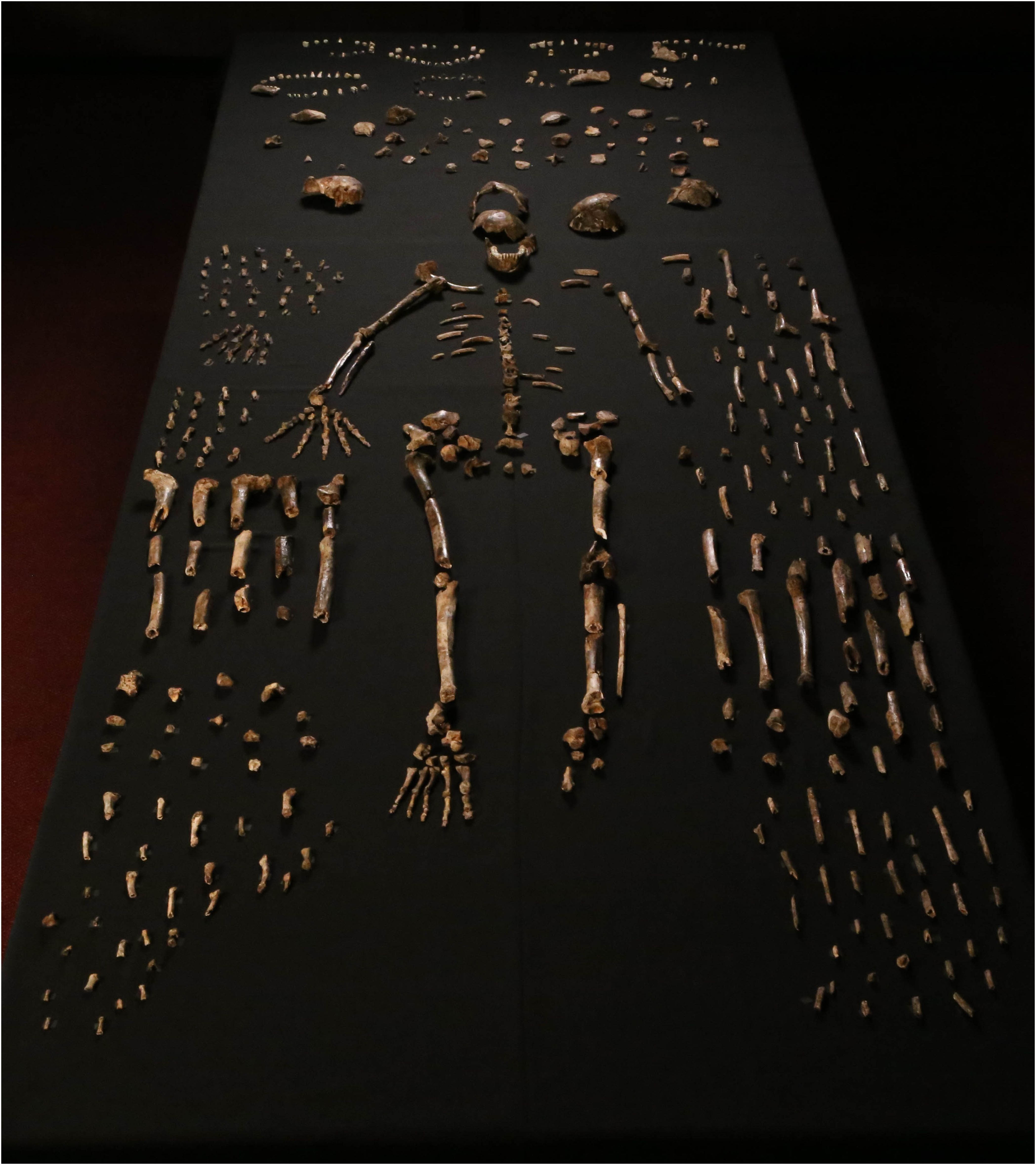
El material fósil descrito en el artículo es realmente notable por varias razones. En primer lugar, la gran cantidad de restos óseos y dentales recuperados (más de 1500, y muchos más esperan ser excavados en el mismo sitio) hace que esta sea "la colección más grande de una sola especie de homínido que se ha descubierto en África hasta ahora"[2]. Un mínimo de 15 personas está representado en el conjunto, las cuales abarcan diferentes grupos de edad, desde bebés hasta adultos mayores. Los restos se encuentran bastante bien conservados, con alguna rotura y evidencia de abrasión superficial, y generalmente están desarticulados excepto en algunos casos notables (p. ej., los huesos de una mano y los de un miembro inferior aún articulados) (fig.1).
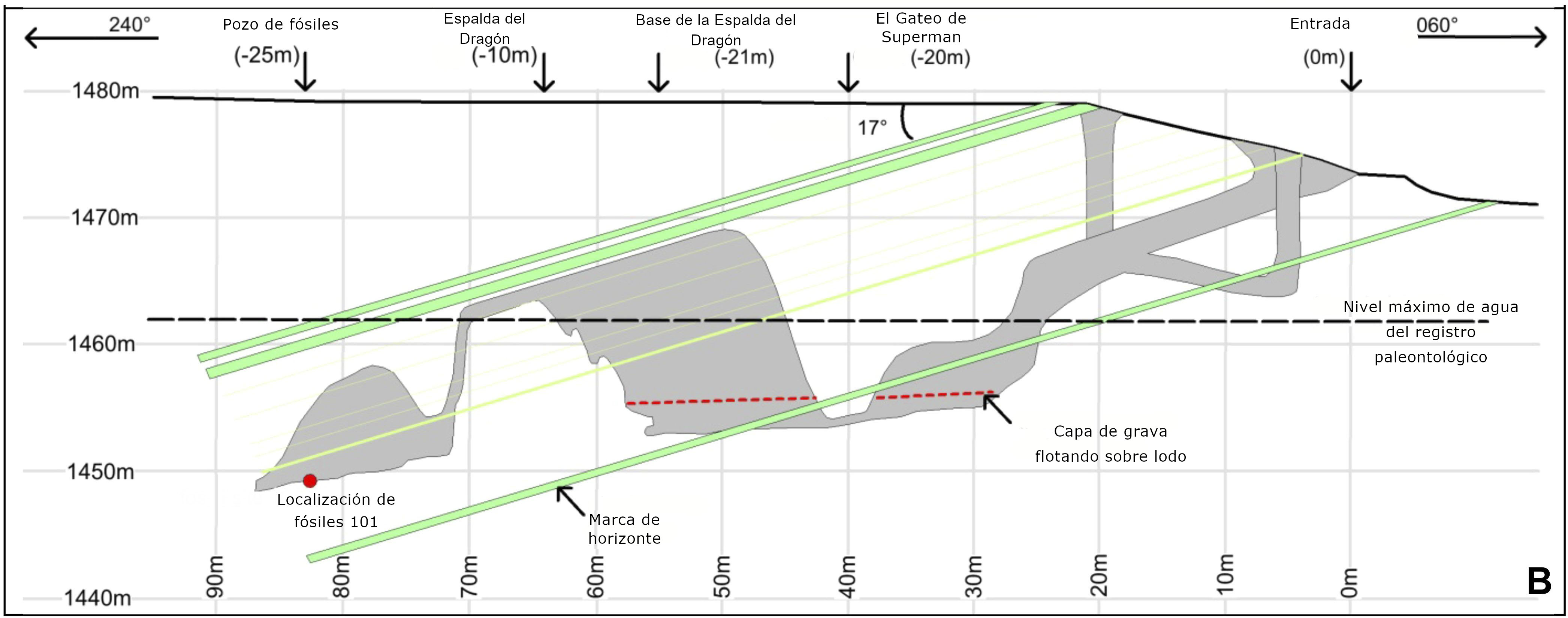
El segundo aspecto que llama la atención en las noticias es la mezcla de rasgos anatómicos similares a humanos y australopitecinos exhibidos en diferentes partes recuperadas (por ejemplo, “manos y pies humanos” y “pelvis similar a australopitecinos” [3]). Finalmente, el contexto en el que se conservan los fósiles también es extremadamente intrigante. Los restos se encontraron en los 20 cm superiores de sedimento fino que cubre el piso de una cámara profunda en un sistema de cuevas (llamado Rising Star) cerca de Johannesburgo (Sudáfrica), actualmente accesible solo a través de pasajes estrechos (hasta 20 cm de diámetro). con caídas verticales de más de 10 m (Fig. 2). No parece que la cámara de la cueva tuviera diferentes accesos en el pasado ni que el agua transportara los restos en la cámara. Además, el conjunto está formado exclusivamente por restos de homínidos, sin restos de otros animales mezclados (con la excepción de los huesos de un búho y algunos dientes de roedor). Esta configuración ha llevado a la especulación de que los cadáveres podrían haber sido desechados intencionalmente en este receso profundo y completamente oscuro, como se analiza en un documento adjunto en eLife (descargable en http://elifesciences.org/content/4/e09561)[4].
Para quienes atesoran el conocimiento, la ciencia y la Palabra de Dios, descubrimientos como este siempre provocan una reacción ambivalente. Estamos emocionados por una nueva evidencia traída del pasado, pero nos entristece verla invariablemente presentada a las masas con un respaldo sin filtros de la evolución darwiniana. Por ejemplo, en el espacio de unos pocos párrafos, un artículo de CNN sobre Homo naledi [5] usa expresiones como "nueva especie de ancestro humano", "una nueva adición a nuestro árbol genealógico" y "nueva especie de pariente humano". El mensaje implícito en este uso del lenguaje es que los humanos no son el resultado de un acto intencional de la creación divina, sino que son solo una de las muchas ramas del árbol de la descendencia común universal de formas ancestrales. Este punto de vista contrasta con el modelo alternativo de Dios creando distintos tipos (incluidos los humanos) que podrían haberse modificado con el tiempo. El lenguaje es una herramienta poderosa que se puede utilizar para promover interpretaciones específicas de datos. Sin embargo, en general, los datos en sí mismos pueden ser compatibles con múltiples hipótesis. El objetivo de este artículo es utilizar el ejemplo de Homo naledi para ilustrar la distinción entre datos e interpretaciones, y discutir algunas de las preguntas que un creacionista bíblico podría tener en relación con este nuevo descubrimiento.
Comprender las limitaciones de los datos
El conjunto de fósiles de Homo naledi es un excelente ejemplo de las limitaciones intrínsecas a la práctica de las ciencias históricas. Solo podemos trabajar con lo que está preservado y disponible, y construir escenarios plausibles basados en la inferencia más lógica, excluyendo hipótesis no respaldadas. Aquí hay algunos ejemplos para ilustrar el punto:
- Acompletamiento. En términos de cantidad de restos fósiles y representación de diferentes partes del cuerpo de un solo sitio, el descubrimiento en la cueva Rising Star es muy impresionante. Sin embargo, incluso en medio de esta abundancia, el registro sigue siendo fragmentario. Por ejemplo, hasta ahora se han recuperado restos de al menos 15 individuos, pero solo están representados cinco cráneos parciales y algunos otros fragmentos craneales. Algunas partes del cráneo no se conservan o están documentadas solo por un espécimen. Dadas estas limitaciones, los rasgos que se presentan como diagnósticos para el cráneo de Homo naledi todavía se sustentan en una base de datos relativamente limitada. El artículo especifica que los diferentes especímenes que conservan la misma parte del cráneo “concuerdan estrechamente en todos los detalles morfológicos”. Sin embargo, también menciona excepciones, en las que las diferencias entre los cráneos se interpretan “en relación con el sexo” [6], que se debe a la variabilidad entre individuos masculinos y femeninos. Desafortunadamente, el documento no proporciona una discusión detallada de las diferencias sugeridas.
- Formas. Es importante comprender la forma en que se obtiene cierta información cuantitativa de los fósiles. Por ejemplo, en la Tabla 1 del artículo [7], se registra la capacidad craneal (una medida del tamaño del cerebro) del Homo naledi como 513 cc. En la sección de métodos del artículo se ofrece una buena explicación de cómo se obtuvo esta cifra. El proceso implicó fusionar dos conjuntos de restos craneales en dos modelos 3D diferentes, interpolar áreas donde no había datos disponibles y luego hacer un promedio entre los dos volúmenes calculados (Fig. 3). Este proceso de cálculo da una indicación clara del tamaño promedio del cerebro que parece representativo en al menos 4 individuos, pero no es un número obtenido de la medición directa de una muestra completa. Otro ejemplo es la estimación de la masa corporal. El trabajo estimó la masa corporal de Homo naledi a partir de huesos de ocho individuos, obteniendo un rango entre 39,7 kg y 55,8 kg [8]. Estos números se calcularon midiendo la amplitud de los huesos del fémur en ubicaciones estándar y aplicando a las mediciones una ecuación de regresión basada en muestras humanas modernas, donde la masa corporal y la amplitud femoral de los individuos humanos se pueden establecer y correlacionar con precisión. Este es un procedimiento estándar, pero está claro que la masa corporal estimada variará si se utilizan diferentes muestras humanas modernas como referencia y dependiendo de las medidas femorales específicas utilizadas. Por ejemplo, utilizando una de las mismas medidas adoptadas en el documento [9], pero con una ecuación de regresión basada en una muestra humana diferente [10] se obtiene un rango entre 33,09 kg y 40,05 kg, que es menor que los valores de masa corporal presentados en el artículo. Además, la suposición de que la correlación entre la amplitud femoral y la masa corporal en los humanos modernos es la misma que en los homínidos fósiles anatómicamente diferentes es obviamente no comprobable y probablemente una aproximación en el mejor de los casos.
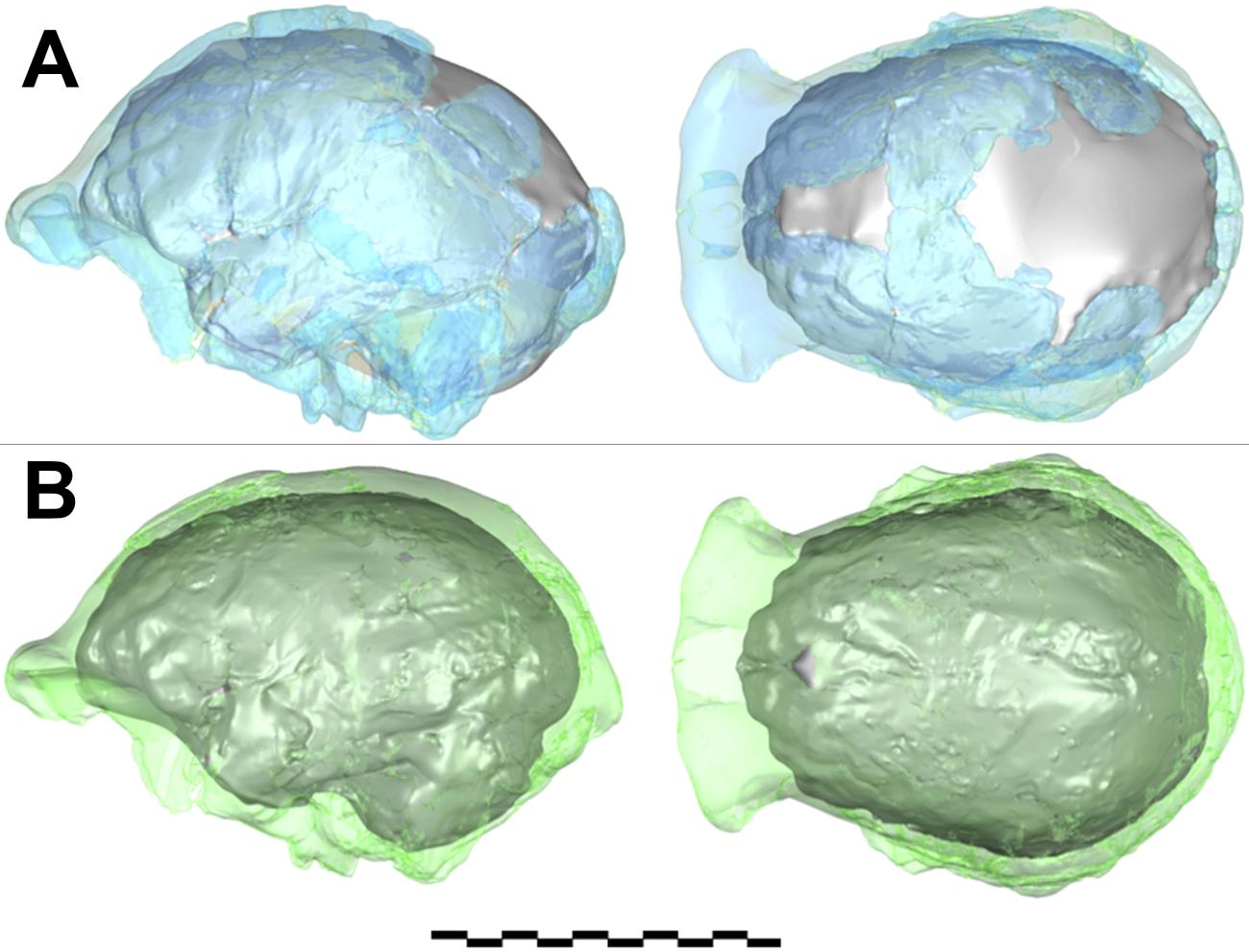
A): compuesto formado a partir de especímenes DH3 y DH4. Este es un cráneo virtual con un un volumen de 465 cm3. La imágen original no tiene medida escala.
B): compuesto formado a partir de especímenes DH1 y DH2. Este es un cráneo virtual más grande calculando una capacidad craneal de 560 cm3. La escala provista es métrica. (Berger et al. 2015 http://elifesciences.org/content/4/e09560)
- Atribución. Debido a que la mayoría de los restos no se encontraron articulados (pero ver una excepción en la Fig. 4), es difícil establecer qué huesos pertenecen al mismo individuo. Paradójicamente, si solo estuviera representado un esqueleto en el conjunto, incluso los restos parciales podrían haberse utilizado para medir directamente las proporciones corporales reales. Tener abundante material disociado permite una mejor comprensión del rango de variabilidad morfológica y de tamaño en una población, pero las reconstrucciones de las proporciones corporales pueden ser más complicadas. Otra posible complicación con los restos disociados es que podría tratarse de una mezcla de dos tipos distintos de homínidos (por ejemplo, uno más parecido a un humano y otro más parecido a un australopitecino) como si pertenecieran a un solo tipo. Los autores del artículo usan una buena ilustración para excluir esta posibilidad. Señalan que "todos los casos en que los elementos se repiten en la muestra, son morfológicamente homogéneos, con una variación consistente en el tamaño corporal y las diferencias de sexo dentro de una sola población" y que "configuraciones morfológicas distintivas", incluidos "rasgos que no se encuentran en especies de homínidos" aún descritas”, son idénticas en las muestras recuperadas. Su conclusión es que “estas consideraciones indican claramente que este material representa una sola especie y no un ensamblaje mixto” [11]. Sin embargo, incluso si este razonamiento es en efecto convincente, la posibilidad teórica de mezclar dos tipos permanece, y fue indicada en los comentarios del descubrimiento por los reconocidos paleoantropólogos Jeffrey Schwartz (Universidad de Pittsburgh) e Ian Tattersall (Museo Americano de Historia Natural) [12].
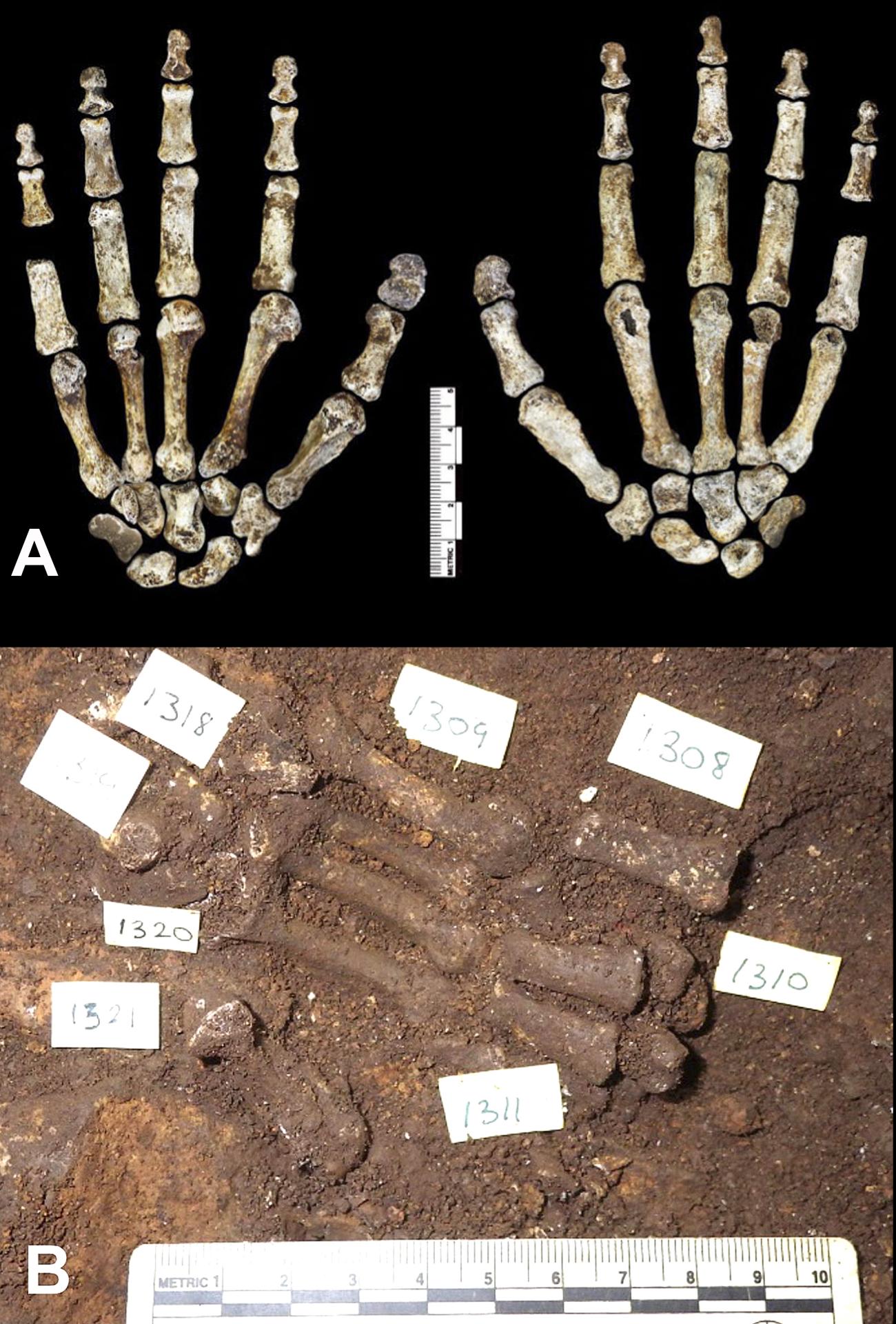
A) Vista desde abajo (en la izquierda) y desde arriba (derecha) Nótese la escala métrica. (Berger et al 2015 http://elifesciences.org/content/4/e09560)
B) Vista de los huesos de las manos durante la excavación. (Dirks et al. 2015 http://elifesciences.org/content/4/e09561#F2)
- Comportamiento. Una demostración final de la brecha entre los datos y las interpretaciones se relaciona con el entorno en el que se encontraron los fósiles de Homo naledi. La lejanía de la cámara de la cueva (Fig. 2) y el hecho de que allí solo se encontraron restos de homínidos ha llevado a afirmar que este es un ejemplo de disposición intencional de cadáveres. Sin embargo, este escenario es solo una posibilidad y no se puede establecer con certeza. Para ser justos con los autores, sus conclusiones reconocen que son posibles otros escenarios: "Tanto el escenario de la mortalidad masiva o la trampa mortal [...] como la hipótesis de eliminación deliberada se consideran interpretaciones plausibles y requieren una investigación adicional", aunque los autores expresan una preferencia por la inferencia de deliberada disposición corporal [13]. Lamentablemente, la cautela y el equilibrio expresados en el artículo no se tradujeron en las declaraciones de los autores a los medios de comunicación, donde se difundieron declaraciones sesgadas como estas: “Acabamos de encontrarnos con otra especie que quizá pensó en su propia mortalidad, y fue a gran riesgo y esfuerzo para disponer de sus muertos en una cámara profunda y remota" [14], “Podemos decir que no se trataba de un grupo social que murió por algún tipo de catástrofe en la cueva” [15]; “Después de eliminar todo lo probable, llegamos a la conclusión de que Homo naledi estaba utilizando esta cámara de forma ritualizada para deshacerse deliberadamente de sus muertos” [16]. Las afirmaciones de comportamiento ritualista, pensamientos sobre la propia mortalidad y la exclusión asertiva del escenario de mortalidad masiva de un grupo social no solo representan una extensión de la evidencia disponible, sino que en el último caso también están en clara contradicción con las conclusiones del artículo.
La agenda del Homo
La palabra Homo tiene un gran significado, porque forma parte del nombre científico de los humanos. Si se coloca un tipo extinto dentro del género Homo, significa que comparte una similitud más cercana con los humanos que con cualquier otro grupo conocido de organismos, incluidos los australopitecinos. Por lo tanto, la elección de un nombre no es solo un tecnicismo trivial, sino que transmite una sensación de cómo les gustaría a los descubridores de un fósil que se percibiera. Para los evolucionistas, algo llamado Homo toma automáticamente cierto lugar en la supuesta secuencia evolutiva de los australopitecinos a los humanos. Para los creacionistas, algo llamado Homo cae automáticamente dentro de las variaciones morfológicas de un cierto tipo creado original. Ambos grupos querrían evitar llamar a Homo algo que no lo es.
Los descubridores de los fósiles de la cueva Raising Star tuvieron que abordar dos preguntas al elegir cómo nombrar los restos: 1) ¿Estos fósiles representan un nuevo tipo? 2) Si es así, ¿a quién se parece más? Al elegir el nombre Homo naledi, respondieron afirmativamente a la primera pregunta y sugirieron humanos como respuesta a la segunda pregunta. Aunque esta elección puede ser válida, es importante comprender que representa una interpretación específica de los datos y que otras interpretaciones pueden sugerir conclusiones diferentes a ambas preguntas. Sobre el aspecto de que se trata de un tipo diferente, por ejemplo, el paleoantropólogo Tim White (Universidad de California, Berkeley) considera, en una etapa temprana de evaluación, que estos restos podrían incluirse dentro de la variabilidad de la especie ya establecida Homo erectus [17].
En la elección de la atribución al género Homo, los autores basan su decisión en aspectos anatómicos similares a los humanos descritos en la extremidad inferior, la mano y los dientes, así como en la forma general del cráneo [18]. Sin embargo, se hace hincapié deliberadamente en estos caracteres, mientras que muchos otros rasgos esqueléticos (por ejemplo, el área pélvica, la región del hombro, la caja torácica) son más parecidos a los australopitales que a los humanos. El tamaño estimado del cerebro, por ejemplo, no solo es pequeño en valor absoluto en comparación con los humanos modernos, sino que también su proporción relativa a la masa corporal estimada (una medida conocida como cociente de encefalización) es aproximadamente la mitad que la de los humanos modernos y similar a la de los australopitecinos [19]. En algunas de las presentaciones de los resultados, parece que los autores del artículo querían enfatizar deliberadamente un vínculo con los humanos en lugar de los australopitecinos. Por ejemplo, el documento afirma que el “H. naledi tiene un rango de masa corporal similar al de poblaciones humanas de cuerpo pequeño” [20], pero una afirmación igualmente cierta habría sido que el rango de masa corporal del H. naledi es similar a la masa corporal estimada para australopitecinos grandes [21]. El énfasis en la eliminación intencional de cadáveres también se enmarca como un argumento de apoyo para resaltar la complejidad del comportamiento que actualmente solo se ve en humanos. En teoría, la atribución de restos fósiles a un género debería basarse en el tratamiento estadístico de diferentes medidas esqueléticas y dentales que agrupan de forma inequívoca especímenes con rasgos anatómicos distintos. En la práctica, el procedimiento se complica por la falta de acuerdo sobre qué rasgos esqueléticos deben considerarse diagnósticos de Homo y por la naturaleza fragmentaria del registro fósil [22].
Evolución humana: ¿Una progresión lineal o un lío enredado?
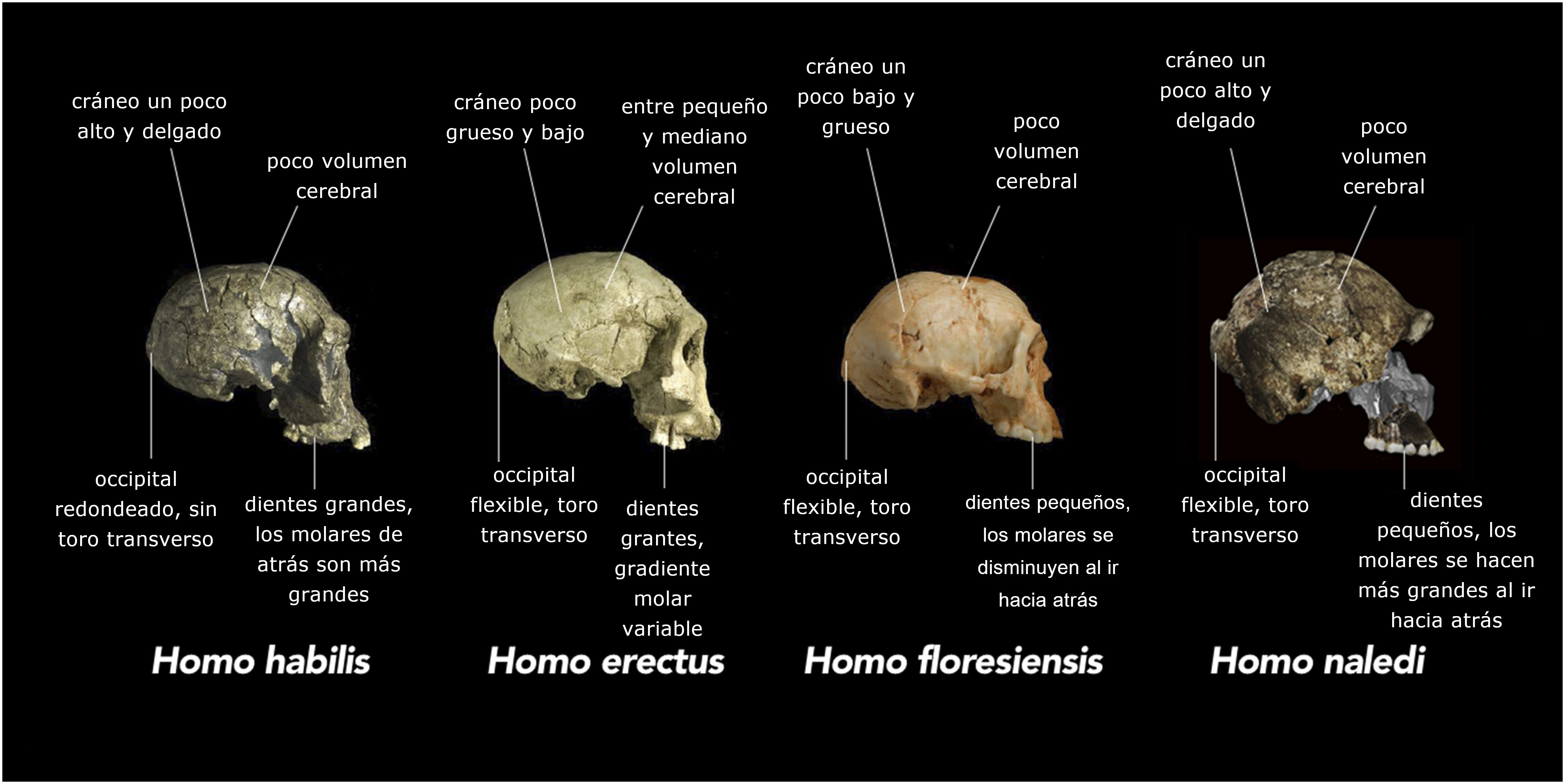
Uno de los aspectos más significativos que surgen del estudio del Homo naledi es su combinación de características anatómicas, algunas más similares a Australopithecus y otras a Homo. Esta distribución en el mosaico de caracteres se ve a nivel de todo el esqueleto, con el tronco, el hombro, la pelvis y la parte proximal del fémur como australopitecinos y los miembros inferiores y la dentición más parecidos a los humanos. Sin embargo, el mosaicismo también es evidente en regiones específicas del esqueleto. Por ejemplo, la mano muestra una morfología general similar al Homo en los huesos de la muñeca, pero dedos relativamente largos y marcadamente curvados (Fig. 4). Los dientes son pequeños, como en los humanos, pero con proporciones de tamaño molar que se encuentran en los australopitecinos. La forma del cráneo es más redondeada, como en el Homo, pero la capacidad craneal y el cociente de encefalización son pequeños, como en los australopitecinos. Se ha documentado una distribución similar en mosaico de caracteres en otros homínidos fósiles, como el Australopithecus sediba [23] y el Homo floresiensis [24].
Además de la mezcla de rasgos esqueléticos observados al nivel de un esqueleto individual, los descubrimientos recientes de Europa del Este han revelado que una cantidad sorprendente de variabilidad morfológica también estaba presente entre diferentes individuos dentro de la misma población de homínidos [25]. Cuando se consideran en conjunto, estos hallazgos presentan un desafío significativo para el escenario clásico de la evolución humana secuencial, popularizado por la icónica procesión de siluetas que se elevan gradualmente desde la postura de simio a la humana. No se puede establecer una trayectoria lineal simple, congruente para el desarrollo de rasgos esqueléticos humanos individuales (Fig. 5). Por el contrario, los complejos funcionales parecidos a los humanos aparecen con una distribución casi "aleatoria" junto con caracteres parecidos a los australopitecinos en diferentes organismos y entre individuos de la misma población. Este cambio de paradigma está siendo reconocido en importantes artículos de revisión. Antón et al., por ejemplo, sostienen que “los ambientes dinámicos favorecieron la experimentación evolutiva y el acoplamiento y desacoplamiento de variables biológicas, lo que impidió cualquier transición simple de Australopithecus a Homo” [26]; también hablan de “intrigante mezcla de rasgos derivados y plesiomórficos y variables biológicas que probablemente caracterizaron la evolución temprana del Homo” [27], es decir, una desconcertante mezcla de rasgos modernos y primitivos desde una perspectiva evolutiva. De manera similar, Schwarz & Tattersall, concluyen que "en contraste con la austera linealidad de Mayr, podemos encontrar que la evolución humana rivalizó con la de otros mamíferos en su experimentación evolutiva y exuberante diversidad" [28]. Palabras como "intrigante", "experimentación", "barajar" y "diversidad exuberante" son fuertes indicadores de una nueva tendencia inesperada y un reconocimiento de lo poco que se sabe realmente con precisión sobre ciertos aspectos del registro fósil de homínidos.
¿Cuál podría ser el origen de esta variabilidad y distribución en mosaico de caracteres en los homínidos fósiles? Como suele ocurrir en las ciencias históricas, solo se pueden hacer sugerencias. Sin embargo, Antón et al. insinúan tres procesos posibles: 1) Plasticidad morfológica y del desarrollo relacionada con las condiciones ambientales. Por ejemplo, la disponibilidad de alimentos y el bajo riesgo de mortalidad pueden prolongar el crecimiento y retrasar la pubertad en los seres humanos, lo que resulta en un tamaño corporal más grande y una maduración más lenta; 2) Hibridación entre diferentes grupos, es decir, la generación de descendientes a partir de individuos pertenecientes a distintos tipos que pueden cruzarse; y 3) Vicarianza, que significa diversificación de grupos como resultado del aislamiento debido a la migración o barreras geográficas [29]. Es interesante darse cuenta de que estos mismos mecanismos funcionarían muy bien también dentro de un marco creacionista, donde se esperaría que los tipos australopitecinos y humanos se diversificaran en una dispersión posterior al diluvio.
Hay un corolario final importante del patrón de distribución en mosaico de rasgos que debe destacarse. Como señalaron deliberadamente Berger et al., debido a las combinaciones no predecibles de caracteres aparentes en diferentes homínidos fósiles "debemos abandonar la expectativa de que cualquier pequeño fragmento de la anatomía pueda proporcionar una visión singular sobre la relación evolutiva de los homínidos fósiles" [30]. En otras palabras, una reconstrucción de supuestas relaciones evolutivas basadas en comparaciones de un solo elemento (por ejemplo, las cúspides de un diente, la forma de una mandíbula, la prominencia de un borde de cejas) conduciría a resultados contradictorios si se dispusiera de esqueletos completos.
Lecciones para aprender
La publicidad dada al descubrimiento del H. naledi ciertamente planteará preguntas sobre el tema de los orígenes humanos. Esta gran oportunidad para una reflexión más profunda sobre el tema será beneficiosa si se pueden evitar los conceptos erróneos. Aquí hay algunos puntos importantes que deben ser considerados por y para aquellos que comparten una perspectiva creacionista:
- ¿Es el Homo naledi el "eslabón perdido" entre los australopitecinos y los humanos? Independientemente de lo que los medios quieran que creamos, la respuesta es no. En primer lugar, los fósiles del Homo naledi no han sido fechados y se desconoce su ubicación estratigráfica. Bien podrían ser un grupo de homínidos relativamente reciente, por lo tanto, irrelevantes en la supuesta aparición evolutiva de Homo. Incluso si se determinó que su posición estratigráfica estaba entre los fósiles de australopitecinos y Homo, el naledi no presenta una disposición transicional de rasgos morfológicos, sino, como hemos visto, una combinación de mosaico, por lo que no ejemplifica una ruta hipotética de cambios secuenciales graduales esperados en la evolución Darwiniana. Aún queda una brecha más significativa en el registro fósil de homínidos, lo que hace que la transición de australopitecinos a humanos sea una interpretación basada en datos muy escasos.
- ¿Qué era naledi? ¿Humano o australopitente? Como se discutió anteriormente, esto es una cuestión de interpretación. Probablemente, los creacionistas se inclinarán a verlo como una variante de tipo australopitecino, especialmente a la luz de su pequeño tamaño cerebral, con adaptaciones para el bipedalismo y modificaciones en la dentición impulsadas por la dieta [31]. La realidad es que simplemente no lo sabemos. Hay varias opciones viables desde una perspectiva creacionista. El H. naledi podría ser un híbrido entre dos tipos, un tipo diferente de primate o una variante impulsada por el medio ambiente, ya sea del tipo australopiteral o del tipo humano.
- ¿Qué no era el naledi? Un nuevo descubrimiento también es una excelente oportunidad para recordar lo que NO se ha descubierto. El H. naledi no es un humano anatómicamente moderno. Hasta ahora, no hemos descubierto restos de humanos de aspecto moderno, excepto hacia la parte superior de la columna geológica. El H. naledi no es un fósil humano gigante. Desafortunadamente, los engaños sobre los descubrimientos de fósiles humanos gigantes continúan siendo populares entre cristianos bien intencionados, pero mal informados [32], y se debe ejercer un gran escepticismo cuando se escuchan noticias sensacionales sobre el tema. No es probable que el H. naledi represente un fósil antediluviano. Su deposición dentro de los sedimentos superficiales de una cueva coloca su existencia al final de la secuencia de eventos geológicos que formaron el paisaje actual.
- ¿Qué lecciones más importantes se pueden aprender de este asombroso descubrimiento? Ciertamente, la lección más instructiva es que todo buscador de conocimiento siempre se beneficiará de una comprensión clara de la diferencia entre datos e interpretaciones. Los creacionistas bíblicos deben ser conscientes de las poderosas presuposiciones filosóficas que operan al presentar nuevos descubrimientos, a menudo arraigados en una cosmovisión evolutiva. Esto puede conducir a un sesgo intencional o no intencional en la presentación de resultados o en la popularización de sus implicaciones. Mientras agudizamos nuestro ojo crítico, debemos continuar trabajando con confianza hacia la construcción de modelos que sean compatibles con una cosmovisión bíblica, manteniendo una actitud humilde en áreas de comprensión incompleta.
Ronny Nalin, PhD
Geoscience Research Institute
Para obtener información actualizada sobre el Homo naledi incluyendo información de documentos publicados en Mayo del 2017 visite nuestro blog en este link.
Texto traducido del inglés por Glendy Noemí Catari
Referencias
[1] Berger, L.R., et al., Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife, 2015. 4.
[2] Ibid., p.3.
[3] Ibid., pp.3,24.
[4] Dirks, P.H., et al., Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife, 2015. 4.
[5] McKenzie, D. and Wende, H., Homo naledi: New Species of Human Ancestor Discovered in South Africa. CNN, September 10, 2015, http://www.cnn.com/2015/09/10/africa/homo-naledi-human-relative-species/.
[6] Berger et al., p.6.
[7] Ibid., p.11.
[8] Ibid., p.18.
[9] AP subtrochanteric breadth, Table 3, p.18 of Berger et al.
[10] Grabowski, M., Hatala, K. G., Jungers, W. L., & Richmond, B. G., Body mass estimates of hominin fossils and the evolution of human body size. Journal of human evolution, 2015. 85, 75-93.
[11] Berger et al., pp.4,5.
[12] As reported in Barras, C., New species of extinct human found in cave may rewrite history. New Scientist, September 10, 2015, https://www.newscientist.com/article/mg22730383-700-new-species-extinct-human-found-in-cave-may-rewrite-history/.
[13] Dirks et al., p.30.
[14] Reported in McKenzie and Wende.
[15] Reported in Johnson, M.A., and Jackson, H., 'Mind Blown': Is Human Ancestor Discovery the Long-Sought Missing Link? NBC News, September 10, 2015, http://www.nbcnews.com/science/science-news/mind-blown-human-ancestor-discovery-long-sought-missing-link-n425406.
[16] Reported in Sample, I., Homo naledi: new species of ancient human discovered, claim scientists, The Guardian, September 10, 2015, http://www.theguardian.com/science/2015/sep/10/new-species-of-ancient-human-discovered-claim-scientists.
[17] Reported in Sample and in Hartley, R., Some bones to pick, Times Live, September 18, 2015, http://www.timeslive.co.za/thetimes/2015/09/18/Some-bones-to-pick.
[18] Berger et al., p.23.
[19] Using the body mass and brain size estimates of Berger et al. and data end equations in Ruff, C.B., Trinkaus, E. and Holliday, T.W. (Body mass and encephalization in Pleistocene Homo. Nature, 1997. 387, 173-176) the estimated encephalization quotient for H. naledi is about 2.4, whereas for modern humans it is about 5.3.
[20] Berger et al., p.18.
[21] Por ejemplo, cuando las mismas ecuaciones y tipos de medidas son utilizadas para calcular la masa corporal en Berger et al. asignadas a AL 333-131 (fémur de Australopithecus afarensis) se obtiene ~61kg, lo cual es una masa coporal mucho mayor que la que se calcula del H. naledi. Al aplicar análisis estadístico de variable múltiple, Grabowski et al. resulta un masa corporal promedio para un macho A. afarensis de 49.5 kg, que de la misma manera excede el cálculo obtenido al aplicarlo a H. naledi.
[22] Ver Schwarz, J.H. and Tattersall, I., Defining the genus Homo. Science, 349/6251, pp.931-932.
[23] Berger, L. R., de Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H. and Kibii, J. M., Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science, 2010. 328/5975, pp.195-204.
[24] Brown, P., Sutikna, T., Morwood, M. J., Soejono, R. P., Saptomo, E. W. and Due, R. A., A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature,2004. 431/7012, pp.1055-1061.
[25] Lordkipanidze, D., de León, M. S. P., Margvelashvili, A., Rak, Y., Rightmire, G. P., Vekua, A. and Zollikofer, C. P., A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 2013. 342/6156, 326-331.
[26] Antón, S. C., Potts, R., and Aiello, L. C., Evolution of early Homo: An integrated biological perspective. Science, 2014. 345/6192, p.1236828-10.
[27] Ibid.
[28] Schwarz and Tattersall, p.932.
[29] Antón et al.
[30] Berger et al., p.23.
[31] Esta es, por ejemplo, la posición adoptada por E. Mitchell en un blog de Respuestas en Génesis en el H. naledi discovery: Is Homo naledi a New Species of Human Ancestor? 12 de Septiembre del 2015, https://answersingenesis.org/human-evolution/homo-naledi-new-species-human-ancestor/
[32] La última invención que me señaló recientemente un amigo, que tenía algunas esperanzas de su credibilidad, se puede encontrar en http://worldnewsdailyreport.com/5-meter-tall-human...